

2.1. Закономірності в нуклеотидному складі днк (правила е.Чаргаффа)
1 |
Сума піримідидових нуклеотидів дорівнює сумі пуринових нуклеотидів: Пурини = Піримідини |
2 |
Вміст тиміну дорівнює вмісту аденіну, а вміст гуаніну дорівнює вмісту цитозину: Т=А, Г=Ц |
3 |
Кількість 6 аміногруп дорівнює кількості 6–кетогруп: Г+Т=А+Ц |
4 |
Відношення
k=
>1–АТ–тип ДНК (вищі рослини, тварини і багато мікроорганізмів) <1–ГЦ–тип ДНК (більшість мікроорганізмів, особливо бактерій і грибів) |
5 |
Відношення
від 0,45 до 2,8 - мікроорганізми від 0,55 до 0,93 - вищі рослини і тварини |
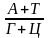 видоспецифічне
і називається коефіцієнтом
специфічності
видоспецифічне
і називається коефіцієнтом
специфічності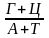 називається фактором
специфічності
називається фактором
специфічності
В молекулі ДНК можна виділити первинну, вторинну і третинну структури. Первинна структура визначається послідовністю нуклеотидів у ланцюгу; вторинна-комплементарністю антипаралельних ланцюгів, з’єднаних водневими зв’язками; третинна-вивчена недостатньо і пов’язана із тримірною просторовою конфігурацією молекул.
Поліморфізм ДНК-здатність подвійної спіралі набувати різних конфігурацій. Поліморфізм залежить від послідовності пар основ, типу пентозного цукру та зовнішніх умов. Рентгеноструктурні дослідження виявили основні типи ДНК: А,- В-, С- і Z- форми (рис.2).

Рис. 2.6. Поліморфізм молекули ДНК (А, В і Z- форми: зліва направо)
В-ДНК-це стандартна уотсон-кріковська структура. Права спираль, з 10 парами основ на виток. Саме в цій формі ДНК існує за фізіологічних умов in vivo.
А-ДНК–також права спіраль, на виток спіралі припадає 11 пар основ, має значний нахил площин пар основ відносно осі спіралі. Реалізується у ДНК лише in vitro за певних умов, далеких від фізіологічних. Проте саме в А-формі існують подвійні спіралі РНК за фізіологічних умов (причиною є заміна дезоксирибози на рибозу). Крім того, ДНК може переходити в А-форму або наближену до неї в комплексах із білками. А-форма виявлена у зневодненому середовищі при високому вмісті калію і натрію. Утворюється при висушуванні волокон В-ДНК.
С-ДНК-має менше основ на один виток спіралі і як слід, інші фізичні характеристики, малодосліджена.
Z-ДНК-ліва спіраль із 12 парами основ. Літера Z свідчить про зигзагоподібну форму цукрофосфатного кістяка ДНК у цій формі (А.Річ). Ця форма зустрічається на ділянках, які збагачені парами Г-Ц. Такі послідовності є в природних ДНК, але перехід у Z-форму відбувається in vitro за умов, які дуже далекі від фізіологічних. Біологічне значення Z-форми залишається не зовсім зрозумілим, хоча знайдено білки, що мають високу спорідненість саме до неї, тобто можуть індукувати В→Z перехід in vivo, а також відіграє істотну роль у процесах рекомбінації і регуляції дії генів.
РНК. Проте ДНК не бере участі в життєдіяльності клітин. Роль посередників у передачі спадкової інформації від ДНК у цитоплазму відіграють рибонуклеїнові кислоти. Взаємовідносини ДНК, РНК і білків можна представити у вигляді схеми:
ДНК→ РНК→Білок
У цьому випадку один з ланцюгів ДНК є матрицею для молекул РНК, що, зокрема, є матрицями синтезу білків або входять до складу рибосом чи переносять амінокислоти.
РНК мають вигляд довгих нерозгалужених полімерних молекул, що складаються з одного ланцюга. Одноланцюгові РНК можуть утворювати подвійні спіралі, якщо різні частини ланцюга мають антипаралельні комплементарні сегменти, пов'язані один з іншим. У частини вірусів РНК є носієм спадкової інформації за відсутності ДНК. Деякі РНК мають каталітичну активність на певні клітинні процеси. РНК-полімер рибонуклеотидів, що складається із фосфорної кислоти, рибози й азотистих основ (аденін, гуанін, цитозин, урацил).
Усі різновиди РНК синтезуються на молекулах ДНК за участю ферментів РНК-полімераз на основі принципу комплементарності. При цьому в синтезованій молекулі аденін ДНК комплементарний урацилу РНК, а гуанін-цитозину (А=У; Г=Ц).
Отже, молекули РНК мають багато спільного із структурою ДНК, але відрізняються низкою ознак: а) вуглеводом є рибоза; б) РНК не містить тиміну, його місце в молекулі займає урацил; в) РНК-одноланцюгова молекула; г) правила Чаргаффа не виконуються.
Типи РНК. На основі розміру, структури і функцій молекул розрізняють три типи рибонуклеїнових кислот (РНК): інформаційна, або матрична (іРНК, або мРНК), транспортна (тРНК) та рибосомальна (рРНК).
іРНК, або мРНК-копія (транскрипт) відповідної ділянки ДНК. Вона служить матрицею для синтезу білкової молекули. Кожні три послідовні основи нуклеотидів іРНК називають кодоном, який кодує одну амінокислоту. Вони несуть закодовану інформацію первинної структури білків у цитоплазму, де прикріплюються до рибосом і реалізують цю інформацію. Інформаційна РНК є матрицею для синтезу поліпептидів (білків), тому її називають також матричною. Матрична РНК є шаблоном, на якому будуються поліпептиди відповідно до закладеної генетичної інформації. Звичайно, вона несе інформацію про синтез тільки однієї молекули білка-це так звана моноцистронна іРНК. Іноді вона містить декілька цистронів, розташованих поряд, для різних білків і відома під назвою поліцистронна іРНК. Інформаційна РНК містить інформацію про порядок розташування амінокислот у синтезованому білку. Розташування амінокислот кодується чіткою послідовністю нуклеотидів у молекулі іРНК (генетичний код). Кожній амінокислоті відповідає свій триплет нуклеотидів (кодон). Молекули іРНК складаються з 300-3000 нуклеотидів. Вони становлять 0,5-3,0 % маси всіх РНК клітини. Інформаційна РНК утворюється в ядрі у вигляді незрілої про-іРНК, яка містить і неінформативні послідовності нуклеотидів-інтрони. В результаті сплайсингу (вирізання інтронних ділянок і зшивання екзотів у кінцевий транскрипт) вона "дозріває" і надходить у цитоплазму, де відразу приєднується до рибосом. При цьому сплайсинг може бути спрямований по різних шляхах-альтернативний сплайсинг-призводить до утворення різних кінцевих продуктів-різних білків. Проте іноді, іРНК може накопичуватися у клітинах, зв'язуватися із спеціальними білками, що "консервують" її, з утворенням інформосом. У такому вигляді інформація може тривалий час зберігатися у клітинах. Поштовхом для їх використання є фізіологічні зміни в клітині, що призводять до активації синтезу білка. Наприклад, в овоциті накопичується багато інформосом, а їх ІРНК починає функціонувати тільки після запліднення.
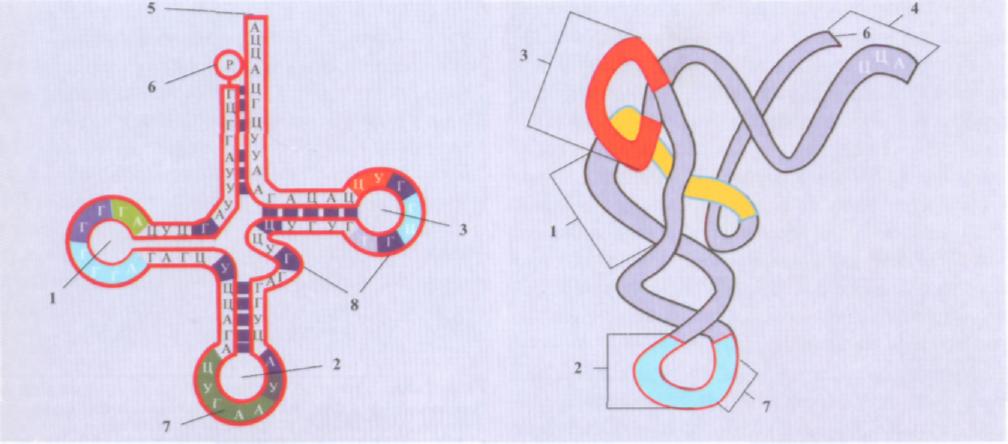
тРНК (ключовий момент системи трансляції)-утворюються на спеціальних генах. тРНК-короткі, однониткові, мають форму листка конюшини завдяки комплементарному сполученню основ на різних ділянках ланцюга, складаються з невеликого числа нуклеотидів-75-90 (рис.2.7). У клітині на долю тРНК приходить 10-20%. Молекули тРНК переносять до місць синтезу білків тільки відповідні їм амінокислоти з цитоплазми. Кожній амінокислоті відповідає своя тРНК внаслідок особливостей нуклеотидної послідовності та просторової структури. Оскільки для кожної амінокислоти є відповідна тРНК, то розпізнавання кодонів іРНК відбувається за допомогою антикодонів молекули тРНК. Антикодон-це три послідовних основи на антикодовій петлі тРНК, які комплементарні основам кодона іРНК:
тРНК: 3´... — А — А — Г — ... 5' — антикодон;
іРНК: 5'... — У — У — Ц — ... 3' — кодон.
Молекули тРНК мають чотири важливі ділянки: а) транспортну; б) антикодон; в) ділянку приєднання фермента; г) ділянку зв'язування з рибосомою.
Один кінець молекули (акцепторний) призначений для приєднання амінокислот, а другий містить антикодон, що складається з трьох нуклеотидів. До транспортної ділянки приєднується специфічна амінокислота. Вона утворена двома комплементарними кінцевими ділянками РНК, 3'-кінець якої складається з семи пар основ. Він довший і формує одноланцюгову ділянку, що закінчується послідовністю ЦЦА з вільною ОН-групою. До цієї групи приєднується амінокислота, що транспортується. Антикодон складається з п'яти нуклеотидів. У центрі-три специфічних рибонуклеотиди (триплет). У період синтезу білка антикодон знаходить відповідний йому кодон на іРНК і тимчасово приєднується до нього водневими зв'язками.
 ис.
2.7. Будова
молекули тРНК:
1-
3
петля (відповідно, 1, 2, 3); 4-акцепторний
кінець; 5-ОН 3'-кінець; 6-5'-кінець; 7-антикодон;
8-модифікаційні нуклеотиди (Пішак
В.П., Бажора Ю.І., Брагін Ш.Б. та ін.,
2004.-С.- 89.)
ис.
2.7. Будова
молекули тРНК:
1-
3
петля (відповідно, 1, 2, 3); 4-акцепторний
кінець; 5-ОН 3'-кінець; 6-5'-кінець; 7-антикодон;
8-модифікаційні нуклеотиди (Пішак
В.П., Бажора Ю.І., Брагін Ш.Б. та ін.,
2004.-С.- 89.)
Ділянка приєднання ферменту-це спеціальна частина молекули тРНК для специфічного зв'язування з ферментом аміноацил-тРНК-синтетазою, що каталізує приєднання амінокислоти до молекули тРНК. Ділянка зв'язування з рибосомою-особлива частина молекули (певна послідовність нуклеотидів) тРНК, що потрібна для прикріплення до рибосоми.
рРНК-складають до 90% усієї РНК клітини, досить стабільні. Утворюються на спеціальних генах ДНК в ядерці. рРНК-велика одноланцюгова розгалужена молекула, що включає 3000-5000 нуклеотидів. У каріоплазмі рРНК і різні білки об’єднуються у співвідношенні 1:1 для утворення малих і великих субодиниць рибосом. Рибосомальна РНК утворює структурний каркас рибосоми, їй належить важлива роль у процесі синтезу білків та забезпечує зв'язування іРНК з рибосомами за допомогою певних послідовностей нуклеотидів. Так встановлюється початок і рамка зчитування інформації з іРНК. Багато білків рибосом виконують не тільки структурну, але й ферментативну функцію.
Таким чином, чотири різновиди нуклеїнових кислот мають багато спільного в будові, але виконують різноманітні функції.