

Способы проведения возбуждения в нервных волокнах. Законы проведения возбуждения
Проведение нервных импульсов по безмякотным и мякотным нервным волокнам принципиально отличается друг от друга. Нервный импульс по безмякотному нервному волокну, не имеющему на своей поверхности электрического изолятора в виде миелина, проводится непрерывно (от точки к точке), вовлекая последовательно в процесс возбуждения все участки волокна (такой способ проведения возбуждения в безмякотных нервных волокнах был впервые описан Германом в 1899 г.).
Р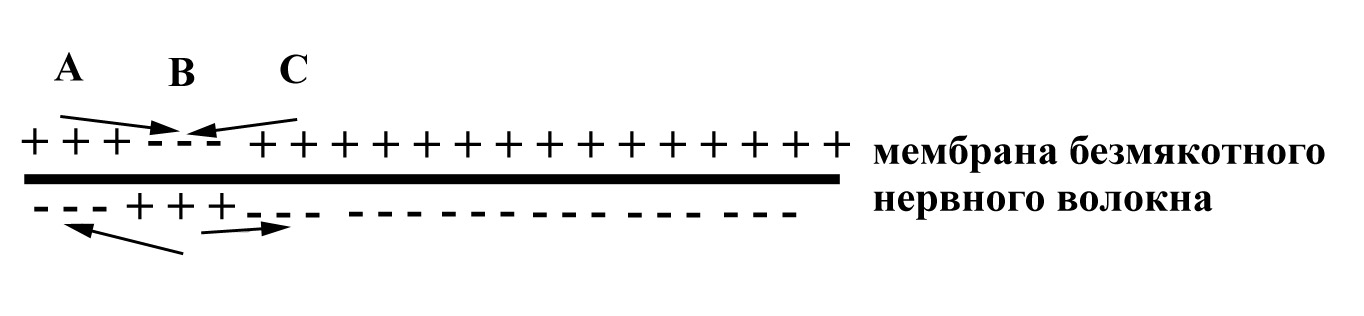 ассмотрим
непрерывное проведение возбуждения по
безмякотному нервному волокну на
примере, представленном на рисунке 9.
Участок А нервного волокна,
изображенный на данном рисунке, только
что был возбужден, его мембрана уже
частично реполяризована после
предыдущего возбуждения, но он
находится пока еще в состоянии
относительной рефрактерности. Этот
участок, будучи ранее возбужденным,
уже вызвал возбуждение соседнего
с ним участка В. В данный момент
(указанный на рисунке) возбужден
участок В: его мембрана заряжена
противоположно по отношению к заряду
мембраны соседних с ним участков –
участка А (только что частично
реполяризованного, но еще остающегося
рефрактерным) и участка С (покоящегося).
Вследствие разных зарядов мембраны
участка В и соседних с ним участков А и
С, между этими участками возникают
местные ионные токи, направленные
от "плюса" к "минусу". Причем,
ток, направленный от участка В к
участку А, будет деполяризующим
по отношению к участку А, поскольку
по аксоплазме волокна в сторону участка
А, заряженного изнутри отрицательно,
движутся положительные ионы. Но такой
деполяризующий ток не способен вызвать
снова возбуждение в участке А, поскольку
участок А еще находится в состоянии
относительной рефрактерности после
только что прошедшего в нем возбуждения.
Ток, от участка А к возбужденному
соседнему участку В, идущий снаружи
нервного волокна (по межклеточной
жидкости), будет деполяризующим по
отношению к участку В, но никаких
изменений в нем в данный момент этот
ток вызвать не может, поскольку
участок В возбужден и абсолютно
рефрактерен. Аналогично, ток, идущий
от покоящегося участка С к возбужденному
в данный момент участку В, хотя и является
деполяризующим по отношению к участку
В, никоим образом не влияет на него,
поскольку участок В временно абсолютно
рефрактерен (его мембрана заряжена
противоположно по отношению к уровню
покоя, что соответствует верхней трети
пика потенциала действия, в момент
которой возбудимая структура абсолютно
рефрактерна). Наконец, ионный ток,
идущий от возбужденного участка В к
покоящемуся участку С по аксоплазме
нервного волокна, будет деполяризующим
по отношению к покоящемуся и
нормально возбудимому участку С и
вызовет начальную пассивную деполяризацию
этого участка, которая повлечет за собой
активную деполяризацию и последующую
генерацию нервного импульса в нем. Затем
возбуждение, аналогично описанной
схеме, от участка С сможет передаться
на следующий за ним покоящийся участок
нервного волокна, но при этом вернуться
назад на предыдущий, ранее бывший
возбужденным, участок не сможет в связи
со временной относительной его
рефрактерностью после возбуждения.
Таким образом, возбуждение в виде
нервного импульса будет проходить
последовательно по всему безмякотному
нервному волокну, захватывая все новые
и новые его участки. Скорость проведения
возбуждения по безмякотному нервному
волокну пропорциональна корню квадратному
из диаметра волокна.
ассмотрим
непрерывное проведение возбуждения по
безмякотному нервному волокну на
примере, представленном на рисунке 9.
Участок А нервного волокна,
изображенный на данном рисунке, только
что был возбужден, его мембрана уже
частично реполяризована после
предыдущего возбуждения, но он
находится пока еще в состоянии
относительной рефрактерности. Этот
участок, будучи ранее возбужденным,
уже вызвал возбуждение соседнего
с ним участка В. В данный момент
(указанный на рисунке) возбужден
участок В: его мембрана заряжена
противоположно по отношению к заряду
мембраны соседних с ним участков –
участка А (только что частично
реполяризованного, но еще остающегося
рефрактерным) и участка С (покоящегося).
Вследствие разных зарядов мембраны
участка В и соседних с ним участков А и
С, между этими участками возникают
местные ионные токи, направленные
от "плюса" к "минусу". Причем,
ток, направленный от участка В к
участку А, будет деполяризующим
по отношению к участку А, поскольку
по аксоплазме волокна в сторону участка
А, заряженного изнутри отрицательно,
движутся положительные ионы. Но такой
деполяризующий ток не способен вызвать
снова возбуждение в участке А, поскольку
участок А еще находится в состоянии
относительной рефрактерности после
только что прошедшего в нем возбуждения.
Ток, от участка А к возбужденному
соседнему участку В, идущий снаружи
нервного волокна (по межклеточной
жидкости), будет деполяризующим по
отношению к участку В, но никаких
изменений в нем в данный момент этот
ток вызвать не может, поскольку
участок В возбужден и абсолютно
рефрактерен. Аналогично, ток, идущий
от покоящегося участка С к возбужденному
в данный момент участку В, хотя и является
деполяризующим по отношению к участку
В, никоим образом не влияет на него,
поскольку участок В временно абсолютно
рефрактерен (его мембрана заряжена
противоположно по отношению к уровню
покоя, что соответствует верхней трети
пика потенциала действия, в момент
которой возбудимая структура абсолютно
рефрактерна). Наконец, ионный ток,
идущий от возбужденного участка В к
покоящемуся участку С по аксоплазме
нервного волокна, будет деполяризующим
по отношению к покоящемуся и
нормально возбудимому участку С и
вызовет начальную пассивную деполяризацию
этого участка, которая повлечет за собой
активную деполяризацию и последующую
генерацию нервного импульса в нем. Затем
возбуждение, аналогично описанной
схеме, от участка С сможет передаться
на следующий за ним покоящийся участок
нервного волокна, но при этом вернуться
назад на предыдущий, ранее бывший
возбужденным, участок не сможет в связи
со временной относительной его
рефрактерностью после возбуждения.
Таким образом, возбуждение в виде
нервного импульса будет проходить
последовательно по всему безмякотному
нервному волокну, захватывая все новые
и новые его участки. Скорость проведения
возбуждения по безмякотному нервному
волокну пропорциональна корню квадратному
из диаметра волокна.
Рис. 9. Схема, отражающая особенности проведения возбуждения по безмякотному нервному волокну. Стрелками показано направление местных ионных токов, возникающих между по-разному заряженными участками мембраны волокна
Возбуждение по мякотным нервным волокнам проводится скачкообразно (сальтаторно): оно возникает только на мембране перехватов Ранвье, не имеющих электрического изолятора (миелина), и, перепрыгивая через межперехватный участок (покрытый миелином), достигает соседнего перехвата Ранвье, инициируя генерацию в нем нового нервного импульса. Так, например, если мембрана перехвата Ранвье, обозначенного буквой В, возбуждена (деполяризована), а мембрана соседнего перехвата (С) нормально поляризована (покоится), то между этими перехватами Ранвье, имеющими различный заряд на мембране будут возникать местные ионные токи (см. рис. 10). Ионный ток, направленный от участка В к участку С, приведет к деполяризации участка С. Подобным образом возбуждение будет постепенно последовательно передаваться на следующие перехваты Ранвье нервного волокна. Ионный же ток от участка В (возбужденного) к участку А (реполяризующемуся после возбуждения) не приведет к генерации в нем нового нервного импульса, потому что участок А находится в состоянии невозбудимости (рефрактерности) после предыдущего потенциала действия, вызвавшего возбуждение в самом участке В.

Рис. 10. Схема, отражающая особенности проведения возбуждения по мякотному нервному волокну. Стрелками показано направление местных ионных токов, возникающих между участками мембраны по-разному заряженных перехватов Ранвье.
Время проведения нервного импульса от одного перехвата Ранвье к соседнему не зависит от длины межперехватного участка и при температуре 24С составляет 0,07 мс. Вместе с тем длина межперехватного участка тем больше, чем больше диаметр волокна. Следовательно, скорость проведения возбуждения по мякотному нервному волокну зависит от его диаметра (пропорциональна диаметру волокна). Впервые предположение о сальтоторном способе проведении возбуждения было высказано Вериго в 1899 г, а экспериментально подтвержден такой способ проведения возбуждения – Като (1924 г.) и Тасаки (1953 г.).
Сальтоторный способ проведения возбуждения по мякотному волокну имеет ряд преимуществ по сравнению с непрерывным проведением в безмякотных:
более быстрый (поскольку возбуждение перепрыгивает через межперехватные участки);
энергетически более экономный (поскольку в состояние возбуждения приходит не вся мембрана волокна, а только лишь перехваты Ранвье, как следствие, потери ионов на единицу длины волокна при возбуждении гораздо меньше таковых в безмякотном волокне, а значит, и меньше тратиться энергии на работу Na+/К+-насоса, восстанавливающего нарушенные трансмембранные градиенты для ионов Na+ и К+ после каждого возбуждения).
Проведение возбуждения как по мякотному, так и по безмякотному нервному волокну требует наличия определенных условий и подчиняется некоторым правилам, получивших название законов проведения возбуждения:
Анатомическая и физиологическая целостность нервного волокна. Для нормального проведения возбуждения по нервному волокну оно должно быть анатомически непрерывным, а электровозбудимые каналы его мембраны должны находиться в состоянии, характерном для покоя (такое состояние обуславливает готовность каналов к возбуждению), т.е. волокно должно быть не только анатомически, но и функционально целостным. Так, например, блокада электровозбудимых натриевых каналов нервного волокна специфическими блокаторами (тетродотоксин, новокаин, лидакаин и др.) на фоне анатомической его целостности приводит к временному блоку проведения нервного импульса через функционально измененный участок.
Изолированное проведение возбуждения по нервным волокнам, составляющим нерв (нерв или нервный ствол – это совокупность (пучок) нервных волокон). Изолированное проведение возбуждения по соседним нервным волокнам в пределах нерва обеспечивается тем, что сопротивление мембран нервных волокон гораздо выше такового межклеточной жидкости, заполняющей пространства между ними. В связи с отмеченным ионные токи, возникающие между по-разному заряженными участками какого-то нервного волокна, шунтируются в межклеточных пространствах, и не способны инициировать возбуждение в соседних волокнах. Невозможность передачи возбуждения от одного нервного волокна к другому в пределах нерва имеет важное физиологическое значение, поскольку нерв, как правило, включает большое количество нервных волокон, имеющих отношение к иннервации различных структур (например, блуждающий нерв имеет отношение к чувствительной и эфферентной иннервации почти всех органов грудной и брюшной полостей). Изолированное проведение возбуждения по нервным волокнам служит основным фактором очень тонкой регуляции деятельности различных органов организма. Нарушение изолированного проведения по нервным волокнам встречается только при развитии определенных патологий в нервной системе. Так, например, при столбнячной инфекции резко снижается сопротивление мембран нервных волокон, и становится возможным эфаптическая передача возбуждения (от одного нервного волокна на соседние в пределах нервного ствола). Отмеченное сопровождается повышением общей возбудимости в нервной системе, нарушением тонкой регуляции деятельности внутренних органов и возникновением целого комплекса соматических и вегетативных расстройств.
Двустороннее проведение возбуждения по нервному волокну. Если в условиях физиологического эксперимента на определенный участок нервного волокна наносить раздражение пороговой силы, то возникающее в данном участке возбуждение проводится двусторонне от места своего возникновения (т.е. к телу нервной клетки (антидромно) и к дистальному концу аксона (ортодромно)). В естественных условиях возбуждение, как правило, возникает в области аксонного холмика (начальный участок аксона в месте его отхождения от тела клетки, лишенный миелиновой оболочки и обладающий в 5-6 раз более высокой возбудимостью по сравнению с мембраной тела и дендритов нейрона). Причем нервный импульс в области аксонного холмика инициируется, как правило, в ответ на тот суммарный потенциал, который возникает в каждый данный момент на мембране тела нейрона в результате прихода информации к соме и дендритам по различным "входам". Возникший в области аксонного холмика нервный импульс распространяется двусторонне – в направлении тела клетки и дистального конца аксона. При этом чаще всего потенциал действия на мембране тела нейрона не возникает, поскольку она отличается значительно более низкой возбудимостью по сравнению с таковой аксона и более высоким входным сопротивлением; поэтому, как правило, имеет место только локальная деполяризация мембраны тела нейрона.