

Вопрос №18История развития представлений о наследственности и изменчивости. Фундаментальные свойства живого.
наследственность и изменчивость как важнейшие свойства любой живой системы обеспечиваются функционированием особого материального субстрата. В ходе исторического развития биологической науки представления о его свойствах, организации и химической природе постоянно расширяются и усложняются.
В 60-х гг. XIX в. основоположник генетики (науки о наследственности и изменчивости) Г. Мендель (1865) высказал первые предположения об организации наследственного материала. На основании результатов своих экспериментов на горохе он пришел к заключению, что наследственный материал дискретен, т.е. представлен отдельными наследственными задатками, отвечающими за развитие определенных признаков организмов. По утверждению Менделя, в наследственном материале организмов, размножающихся половым путем, развитие отдельного признака обеспечивается парой аллельных задатков, пришедших с половыми клетками от обоих родителей. При образовании гамет в каждую из них попадает лишь один из пары аллельных задатков, поэтому гаметы всегда «чисты». В 1909 г. В. Иогансен назвал «наследственные задатки» Менделя генами. Наследственность – свойство клеток или организмов в процессе самовоспроизведения передавать новому поколению способность к определенному типу обмена веществ и индивидуального развития, в ходе которого у них формируется общие признаки и свойства данного типа клеток и видов организмов, а также некоторые индивидуальные особенности родителей. Изменчивость – свойство живых систем приобретать изменения и существовать в различных вариантах. Несмотря на то, что по своим результатам наследственность и изменчивость разнонаправлены, в живой природе эти два фундаментальных свойства образуют неразрывное единство, чем достигается одновременно сохранение в процессе эволюции имеющихся биологически целесообразных качеств и возникновение новых, делающих возможным существование жизни в разнообразных условиях.
Билет №19. Законы Менделя. Цитологические основы универсальности законов. Менделирующие признаки человека. Примеры. Промежуточный тип наследования. Анализирующий тип скрещивания и его значение.
Открыл законы в 1865. 1-ый закон(единообразия/доминирования): при моногибридном скрещивании 2х гомозиготных организмов всё первое поколение гибридов оказывается единоообразным. (левая половина рисунка) А-ген жёлтого цвета горошин, а-ген зелёного цвета, всё первое поколение-жёлтого цвета.
![]()
2-ой закон(расщепления): при скрещивании 2х особей, потомков 1-ого поколения, в следующем поколении наблюдается расщепление по фенотипу 3:1, по генотипу-1:2:1(правая часть рисунка). Фенотип F2(по порядку): жёлт,жёлт,жёлт,зел.
3-ий закон(независимого наследования признаков): при скрещивании 2х особей, различающихся по 2м и более парам альтернативных признаков, расщепление по каждой паре признаков идёт независимо от расщепления по другим парам признаков.
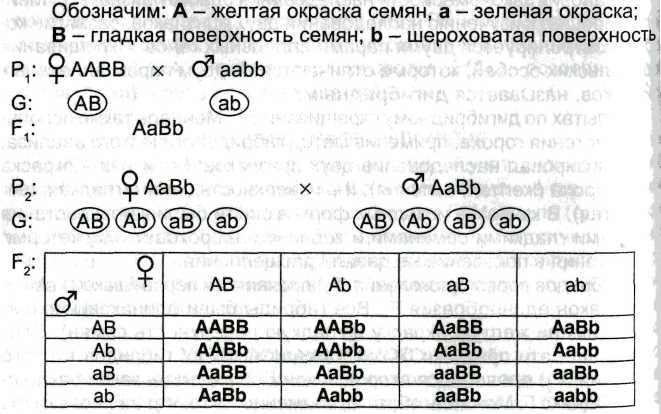
Наблюдаем расщепление по фенотипу: 9:3:3:1 (жёлт.,глад.-9; жёлт.,морщин.-3; зел.,глад.-3; зел.,морщин.-1); жёлтые:зелёные=12:4=3:1; гладкие:морщинистые=12:4=3:1. На рисунке-решётка Пеннета(она позволяет легче и нагляднее представить генотипы, получающиеся при скрещивании родительских гамет.)
Альтернативные признаки-взаимоисключающие признаки(т.е. либо жёлтый цвет, либо зелёный), имеющие контрастное проявление.
Цитологические основы законов Менделя базируются на:
Парности хромосом (парности генов, обусловливающих развитие какого-либо признака);
Особенностях мейоза (процессах, происходящих в мейозе, которые обеспечивают независимое расхождение хромосом с находящимися в них генами к разным полюсам клетки, а затем и в разные гаметы);
Особенностях процесса оплодотворения(случайной комбинации хромосом, несущих по одному гену из каждой аллельной пары).
Признаки, наследование которых подчиняется законам Менделя, называются менделирующими. Примеры: отсутствие пигментации(альбинизм) (рецессивный ген отвечает за альбинизм); цвет глаз; характер волос (курчавые или прямые); I,II,III группы крови; свёртываемость крови. Наследование генов, обусловливающих развитие дальтонизма, гемофилии, тоже подчиняются законам М.
В гетерозиготном состоянии доминантный ген не всегда подавляет проявление рецессивного, и тогда признак у гибридов F1 имеет промежуточный характер. Это и называется промежуточным типом наследования. Наблюдается при наследовании цвета ночной красавицы. Р: АА*аа (красный цвет, белый цвет); G: А, а; F1: Аа(розовый цвет).
Чаще всего по фенотипу нельзя судить о генотипе особи. Лишь в одном случае фенотипу соответсвует строго определённый генотип-в случае, когда особь гомозиготна по рецессивному признаку. Поэтому для того, чтобы определить гомозиготна или гетерозиготна по какому-то признаку исследуемая особь, проводят анализирующее скрещивание, т.е. скрещивание с рецессивной гомозиготной особью.
P: А- * аа; F1: Аа или АА: Аа. Если всё полученное потомство единообразно, то исследуемая особь-гомозиготна, если нет-гетерозиготна.
Билет №20. Аллельные гены. Наследование признаков при взаимодействии аллельных генов. Примеры. Множественный аллелизм. Механизм возникновения.
У организма 1ого вида каждый ген расположен в определённом участке определённой хромосомы. Это место-локус гена. Гены, расположенные в одинаковых локусах гомологичных хромосом и отвечающие за развитие вариантов одного признака, называются аллельными. Если аллельные гены одинаковы по структуре и, следовательно, оба отвечают за определённое проявление признака, то такое состояние является гомозиготным, если не одинаковы-гетерозиготным.
Виды взаимодействия аллельных генов:
1.Полное доминирование (один ген полностью подавляет проявление другого гена. По 1ому закону М.) Пр: нормальная форма эритроцитов над серповидной формой эритроцитов(серповидноклеточная анемия), карий цвет глаз над голубым, нормальное восприятие цветов над дальтонизмом.
2.Неполное доминирование (на примере ночной красавицы)
3.Кодоминирование (отсутствие доминантных и рецессивных отношений между генами) Пр: IV группа крови (ни IA, ни IB не является доминантным геном)
4.Межаллельная комплементация (возможность формирования нормального признака D у организмов, гетерозиготным по 2м мутантным аллелям гена D (D’D’’)
5.Аллельное исключение (в результате инактивации одной из Х-хромосом у особей гомогаметного пола, которая способствует тому, что в разных клетках организма, мозаичных по функционирующей хромосоме, фенотипически проявляется либо материнская, либо отцовская аллель гена) Пр: мозаицизм женского организма по наличию или отсутствию нормальных потовых желёз в коже, обусловленный экспрессией нормального или мутантного аллелей гена Х-хромосомы.
Существует понятие сверхдоминирования- явления преимущества гетерозиготных особей над гомозиготными особями по доминантному гену.
Один из характерных примеров сверхдоминирвания является повышенная частота аллеля гена серповидноклеточной анемии в популяциях человека, живущих в условиях высокой вероятности заражения малярией. Мутантный аллель защищает организм от заболевания малярией. Гомозиготы по нормальному аллелю могут заболеть малярией и погибнуть, гомозиготы по мутантному аллелю - с высокой вероятностью гибнут от анемии. Гетрозиготы по этому гену не болеют серповидновлеточной анемией и устойчивы к малярии.
Множественный аллелизм — это существование в популяции более двух аллелей данного гена. В популяции оказываются не два аллельных гена, а несколько. Возникают в результате разных мутаций гена, занимающего определённый локус. Гены множественных аллелей взаимодействуют между собой различным образом.
В популяциях диплоидных организмов обычно существует множество аллелей для каждого гена. Это следует из сложной структуры гена — замена любого из нуклеотидов или иные мутации приводят к появлению новых аллелей. Примером множественного аллелизма может служит система групп крови АВО, открытая австрийским ученым К. Ландштейнером в 1900 г.
Билет №21. Неаллельные гены. Наследование признаков при взаимодействии неаллельных генов. Примеры.
В зависимости от характера участия неаллельных генов в формировании того или иного признака различают несколько видов их взаимодействия.
Виды взаимодействия неаллельных генов:
Комплементарность (для проявления признака нужна совокупность неаллельных доминантных генов: присутствие в генотипе доминантных аллелей обоих неаллельных генов обеспечивает развитие признака, чего не происходит при отсутствии одного из них в доминантном состоянии; возможна ситуация, когда каждый из неаллельных генов в отсутствие доминантного аллеля другого обеспечивает развитие определённого варианта признака, а вместе они формируют его новый вариант (форма гребня у петухов)). Примеры: каштановый цвет волос, нормальный слух, мужской пол, оттенки цвета глаз.
Полимерия(Совместное действие полигенов (системы неаллельных генов, одинаково влияющих на формирование определённого признака) обусловливает различную экспрессивность-степень выраженности признака, зависящую от дозы соответствующих аллелей) Примеры: пигментация кожи, рост, вес.
Эпистаз (один из генов (В) выражается фенотипически лишь при отсутствии в генотипе определённого аллеля другого гена (А). В его присутсвии действие гена В не проявляется.). Эпистаз бывает:
Доминантный (доминантный аллель одного гена (А) препятствует проявлению аллелей другого гена (В или b)) Примеры:окраска плодов тыквы, цвет оперения кур породы леггорн
Рецессивный (ген, определяющий какой-то признак (В) не проявляется, если особь гомозиготна по рецессивному аллелю другого гена(т.е. аа) Примеры: «Бомбейский феномен», рождение альбиносов у негров.
Эффект положения (это взаимодействие, обусловленное местом положения гена в системе генотипа: непосредственной окружение, в котором находится ген, может сказываться на характере его экспрессии)
Билет № 22. Генетические основы существования групп крови в системе АВ0. Наследование групп крови. Наследование резус- фактора. Резус- конфликт. Основные принципы применения гемотрансфузии в медицине.
За формирование этого признака (группы крови) отвечает ген I. Он обеспечивает синтез антигенов 0, А и В, фиксирующихся на поверхности эритроцитов. Если IoIo, то первая группа (на поверхности эритроцитов антиген 0, а в плазме антитела : антиА и антиВ ); если IoIA или IAIA-вторая (антигены 0, А; антиВ);IoIB или IBIB- третья (антиген 0, В; антиА); IAIB-четвертая (антигены А,В; антител в плазме нет). Наследование группы крови является примером доминирования одного из аллелей в гетерозиготном генотипе: IB, IA доминируют над Io. Четвёртая группа крови наследуется в результате взаимодействия аллелей IA и IB, которое называется кодоминирование.
Но группа крови является сложным признаком, так как он есть результат взаимодействия нескольких пар неаллельных генов, а именно генов I и H. Ген I обеспечивает синтез антигенов А и В. Однако для синтеза антигенов А и В необходимо образование неспецифического вещества-предшественника, которое детерминируется геном Н, находящимся в доминантном состоянии. У гомозигот по рецессивному аллелю hh даже при наличии в генотипе доминантных аллелей гена IA или IB антигены не синтезируются, и фенотипически эти люди имеют первую группу крови (рецессивный эпистаз). Это и есть «Бомбейский феномен»(когда у женщины с первой группой родился ребёнок с четвёртой).
Говоря о резус-факторе, имеется в виду антиген D, фиксирующийся на поверхности эритроцитов. Если он есть, то резус-фактор-положительный, если нет-отрицательный. Ген, отвечающий за синтез антигена D, доминирует над геном, подавляющим синтез.
Если у женщины отрицательный резус-фактор, а у мужчины-положительный, то скорее всего у ребёнка будет резус-положительный, тогда возникает резус-конфликт: организм ребёнка содержит антиген D, а в организме матери его нет, следовательно, организм матери будет вырабатывать антитела на неизвестный антиген (D)/
Небольшие порции крови можно переливать по следующей схеме:
Первую группу можно переливать людям с любой группой крови(людей с первой группой называют универсальными донорами);
Вторую-только второй и четвёртой;
Третью-только третьей и четвёртой;
Четвёртую-только четвёртой.
Таким образом, люди с четвёртой-универсальные реципиенты.
Принципы гемотрансфузии основаны на возможности склеивания эритроцитов реципиентов плазмой доноров. Оно происходит, если встречаются одноимённые антигены и антитела, например, антиген А и антитело А (антиА).
Билет №23. Генотип как единое целое. Ядерная наследственность. Закономерности наследования внеядерных генов. Цитоплазматическая наследственность у про- и эукариот.
Генотип-система взаимодействующих генов в соматической клетки особи. Генотип вместе с внешними факторами определяют фенотип(совокупность всех признаков) организма. Непрерывность существования и историческое развитие живой природы обусловлены двумя фундаментальными свойствами жизни: наследственностью и изменчивостью. Наследственность-свойство живых организмов передавать из поколения в поколение наследственную информацию о развитии организма. Наследственный материал (гены), главным образом, содержится в хромосомах, в ядре, и при делении клетки наследственный материал равномерно распределяется между дочерними клетками.
В начале 20 века были обнаружены факты, которые доказывали наличие в клетках внехромосомного наследственного материала, располагающегося в различных цитоплазматических структурах и определяющего особую цитоплазматическую наследственность (К. Коррес, 1908) Некоторое количество наследственного материала содержится в виде колец ДНК в пластидах и митохондриях, а также в других внеядерных структурах. Цитоплазматические гены не подчиняются менделеевским законам. В связи с тем. Что организм, образуемый в результате оплодотворения, получает цитоплазматические структуры главным образом с яйцеклеткой, цитоплазматическое наследование признаков осуществляется по материнской линии. Такой тип наследования был описан Корренсом в отношении признака пёстрых листьев у некоторых растений.
Билет №24. Типы наследования признаков: независимое, сцепленное с полом, аутосомное, голандрическое, моногенное, полигенное. Примеры.
1.Независимое (разные признаки, определяемые неаллельными генами, передаются потомкам независимо друг от друга и комбинируются у них во всевозможных сочетаниях - третий закон Менделя) Примеры: касается всех признаков, определяемых неаллельными генами,, располагающимися в негомологичных хромосомах.
2.Сцепленное с полом(Х- и У-хромосомы, кроме генов, определяющих пол, содержат гены, не имеющие отношения к полу. Признаки, определяемые этими генами, называются признаками, сцепленными с полом. Иными словами, эти признаки наследуются с полом). Бывают Х-сцепленные признаки, проявляющиеся чаще всего у мужчин и У-сцепленные, проявляющиеся только у мужчин.
3.Голандрическое наследование (наследование признаков, обусловленных генами, содержащимися в У-хромосоме) Примеры: гипертрихоз мочки уха, индактилия (сросшиеся 2-ой и 3-ий пальцы на ноге).
4.Х-сцепленное. Примеры: гемофилия, дальтонизм, отсутствие потовых желёз, атрофия зрительного нерва.
5.Аутосомное (наследование признаков, обусловленных генами, находящимися в соматических(неполовых) клетках)Примеры: цвет глаз, форма ушей, прочность зубов и т.д. и т.п.
6.Моногенное (на формирование признака влияет один ген)Примеры: любой простой признак.
7.Полигенное (на формирование влияют несколько генов) Примеры: описаны при разборе видов взаимодействия аллельных и неаллельных генов.
Билет № 25. Хромосомная теория наследственности. Эксперименты Моргана, доказывающие явление сцепленного наследования и причины нарушения сцепления. Понятие генетических карт хромосом.
Хромосомная теория(1911-1926):
1.Гены расположены в хромосоме в линейной последовательности;
2.Каждый ген занимает определённый локус. Аллельные гены расположены в одинаковых локусах гомологичных хромосом.
3.Гены одной хромосомы наследуются совместно, образую группу сцепления.
4.Число групп сцепления соответствует гаплоидному набору хромосом и постоянно для каждого вида (у женщин-23, у мужчин 24(+У)).
5.Сцепление может нарушаться в результате кроссинговера за счёт образования рекомбинантных хромосом.
6.Частота кроссинговера зависит прежде всего от расстояния между генами: чем оно больше, тем чаще между ними происходит кроссинговер.
Опыты Моргана. Если скрестить мушку дрозофилу, имеющую серое тело и нормальные крылья (самка), с мушкой, обладающей тёмной окраской и зачаточными (короткими) крыльями (самец), то в первом поколении гибридов все мухи будут серыми с нормальными крыльями. Это гетерозиготы по двум парам аллельных генов, причём ген, определяющий серую окраску брюшка, доминирует над тёмной окраской, а ген, обуславливающий развитие нормальных крыльев, - доминирует над геном недоразвитых крыльев.
При анализирующем скрещивании гибрида F1 с гомозиготной рецессивной дрозофилой (чёрное тело и короткие крылья) подавляющее большинство потомков F2 будет сходно с родительскими формами.
Это происходит потому, что гены, отвечающие за серое тело и нормальные крылья - сцепленные гены, также как и гены, отвечающие за тёмное тело и короткие крылья, т.е. они находятся в одной хромосоме. наследование сцепленных генов называют - сцепленное наследование.
Сцепление может нарушаться. Это доказывают особи, имеющие серое тело и короткие крылья; чёрное тело и длинные крылья, т. е. если бы сцепление не нарушалось, то этих особей бы не существовало, однако они есть. Это происходит в результате кроссинговера, который и нарушает сцепленность этих генов.
Генетические карты хромосом. Это схемы относительного расположения сцепленных между собой наследственных факторов — генов. Г.к.х. отображают реально существующий линейный порядок размещения генов в хромосомах и важны как в теоретических исследованиях, так и при проведении селекционной работы, т.к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. Имея Г. к. х., можно по наследованию «сигнального» гена, тесно сцепленного с изучаемым, контролировать передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; например, ген, определяющий эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения.
Билет № 26. Роль наследственных и средовых факторов в определении половой принадлежности организма. Эпигамное, прогамное и сигамное определение пола.
Пол —совокупность свойств, обеспечивающих воспроизведение потомства и передачу ему наследственной информации. В зависимости от значимости этих свойств различают первичные и вторичные половые признаки. Первичные-мофофизиологические особенности организма, обеспечивающие образование гамет, сближение и соединение их в процессе оплодотворения. (наружные и внутренние органы размножения). Вторичные – отличительные особенности особей одного пола от другого, не связанные непосредственно с образованием гамет, спариванием и оплодотворением, но играющие важную роль в половом размножении (обнаружение и привлечение партнёра)(например, половое поведение). Их развитие контролируется гормонами, синтезируемыми первичными половыми органами.
На формирование признаков пола влияет не только наследственная программа, заключённая в генотипе, но и условия окружающей среды. (но у высших организмов значение среды в определении пола, как правило, невелико).
К наследственным факторам определения половой принадлежности относятся: 1.Сочетание половых хромосом, возникающее в зиготе при оплодотворении;
2.Баланс женских и мужских генов - детерминаторов пола (гипотеза генного баланса Бриджеса, согласно которой организм изначально бисексуален, т.е. несёт в себе задатки и мужского, и женского полов. Развитие признаков одного из них определяется соотношением женских и мужских гормонов.) Примеры: гены У-хромосомы, определяющие детерминацию мужского пола, в том числе и ген, определяющий дифференцировку половых желёз по мужскому типу, синтезирующих тестостерон+ген Х-хромосомы, контролирующий синтез белка-рецептора, обеспечивающий проникновение тестостерона в клетки тканей-мишеней(т.е. при проникновении тестостерона в ткани половых желёз(ткани-мишени) из них формируются именно мужские половые органы ). При мутации последнего гена наблюдается явление тестикулярной феминизации (синдром Морриса): кариотип ХУ, но особь внешне более сходна с женщиной. Такие гены содержатся не только в половых хромосомах (главным образом, конечно, в них), но и в аутосомах. Примеры: ген-трансформатор(у дрозофиллы), который в рецессивном гомозиготном состоянии превращает женские зиготы ХХ в фенотипических стерильных самцов, самки гетерозиготные и гомозиготные по доминантному аллелю гена – фенотипически плодовитые самки.
Приведённые примеры доказывают, что пол организмов как признак развивается на основе полученной от родителей наследственной информации и находится под контролем группы взаимодействующих генов, расположенных как в половых, так и в аутосомах.
Известны примеры, когда в определённых условиях развития зиготы происходило переопределение пола на противоположный, несмотря на имеющуюся комбинацию хромосом в зиготе. Примеры: морской кольчатый червь бонеллия, крокодилы (подробное описание ниже)- переопределение пола в эмбриогенезе; рыбки Labroides dimidiatus, обитающие в Тихом океане(они живут стайками, состоящими из одного самца и самок, при чём каждый член такой семьи, кроме самца, находится в состоянии стресса, причина которого-самец. Самки испытывают разные степени стресса. После гибели самца альфа-самка, которая сбрасывала напряжение, превращается в плодовитого самца.) Это пример переопределения пола в онтогенезе. У человека угнетение эндокринной функции яичников, возникающее после прекращения гаметогенеза(в определённом возрасте у женщин), приводит к проявлению некоторых вторичных признаков мужского пола, например росту волос на лице.
У организмов разных видов относительное значение генетических и средовых факторов неодинаково: у одних определяющим фактором является среда, у других-наследственная программа.
Пол особи может определяться: а) до оплодотворения яйцеклетки сперматозоидом (прогамное определение пола); б) в момент оплодотворения (сигамное определение пола);
в) после оплодотворения (эпигамное определение пола).
Прогамное определение. До оплодотворения пол определяется у некоторых организмов в результате разделения яйцеклеток на быстро и медленно растущие. Первые (более крупные) после слияния с мужской гаметой дают самок, а вторые (мелкие) — самцов. У коловраток, способных размножаться помимо обычного полового размножения с оплодотворением, партеногенетически, часть партеногенетических яйцеклеток во время развития лишается половины хромосом. Из таких яиц развиваются самцы, а остальная часть дает начало самкам. Эпигамное . У морского кольчатого червя бонеллия определение пола происходит в процессе онтогенеза: если личинка садится на дно, из нее развивается самка, а если прикрепляется к хоботку взрослой самки, то самец (в данном случае наблюдается влияние внешних условий на формирование пола). В том числе, на определение пола может влиять температура окружающей среды, что характерно для большинства крокодилов. Сигамное. У подавляющего же большинства эукариот пол закладывается в момент оплодотворения и определяется генотипически хромосомным набором, который зигота получает от родителей. Клетки мужских и женских особей животных организмов различаются по паре хромосом. Эту пару называют половыми хромосомами (гетеросомами) в противоположность остальным — аутосомам.
Билет№ 27. Генетика пола. Гомогаметный и гетерогаметный пол. Доказательства генетического определения признаков пола. Хромосомное определение пола у человека и различных организмов.
Различают гомо- и гетерогаметный пол. Представители гомогаметного пола образуют один вид гамет, а гетерогаметный-2. У гомогаметног пола, имеющего одинаковые ХХ хромосомы, все гаметы несут гаплоидный набор хромосом плюс одну Х-хромосому. У гетерогаметного пола в кариотипе кроме аутосом содержатся 2 разные или только одна половая хромосома (ХУ и Х0). Его представители образуют 2 вида гамет, различающихся по гетерохромосомам : Х и У или Х и 0. Доказательства генетического определения признаков пола – в билете № 26 плюс ниже.
В
зависимости от сочетания X-
и Y-хромосом
у
![]() и
и
![]() организмов
различают 5 типов хромосомного определения
пола:
1)
XX,
ХО
(O
обозначает отсутствие хромосом)
встречается у видов Protenor
(насекомые);
2)
XX,
XY
— он характерен, например, для дрозофилы,
млекопитающих (в том числе и для
человека);
3)
XY,
XX
— этот тип определения пола характерен
для бабочек, птиц, рептилий;
4)
ХО,
XX
— наблюдается у тли;
5) гапло-диплоидный тип (
2n,
n)
встречается, например, у пчел: самцы
развиваются из неоплодотворенных
гаплоидных яйцеклеток, самки — из
оплодотворенных диплоидных.
Конкретные механизмы, связывающие
развитие мужского или женского пола с
определенным сочетанием половых хромосом
у разных организмов различен. У человека,
например, пол определяется наличием
Y-xpoмосомы:
в ней есть ТДФ-ген, он кодирует тестикул
- детерминирующий фактор, который
определяет развитие мужского пола.
У дрозофилы же в Y-хромосоме
находится ген фертильности, ответственный
за плодовитость самца, а пол определяется
баллансом числа Х-хромосом и числа
наборов аутосом (это называется половой
индекс)(обычный диплоидный организм
содержит, соответственно, два набора
аутосом). В Х-хромосомах расположены
гены, определяющие развитие по пути
самки, а в аутосомах — по пути самца.
Если отношение количества Х-хромосом
к количеству наборов аутосом равно
0,5(т.е. ХУ: 2 набора аутосом) то развивается
самец, а если — 1(ХХ(2Х): 2 набора аутосом),
то самка.
Помимо нормальных самцов и самок иногда
появляются интерсексы
— особи, по своим половым признакам
занимающие промежуточное положение
между мужским и женским полом (не путать
с гермафродитами!). Это может быть вызвано
как анеуплоидией по половым хромосомам
в гаметах, так и различными нарушениями
(например, гормональными) в процессе
дифференцировки пола.
У большинства
организмов среди потомков получается
50 % самцов и 50 % самок, так как в норме
расщепление по полу подчиняется законам
обычного моногибридного скрещивания
между гомо- и гетерозиготой, для гибридов
которого характерно расщепление в
соотношении 1:1.
организмов
различают 5 типов хромосомного определения
пола:
1)
XX,
ХО
(O
обозначает отсутствие хромосом)
встречается у видов Protenor
(насекомые);
2)
XX,
XY
— он характерен, например, для дрозофилы,
млекопитающих (в том числе и для
человека);
3)
XY,
XX
— этот тип определения пола характерен
для бабочек, птиц, рептилий;
4)
ХО,
XX
— наблюдается у тли;
5) гапло-диплоидный тип (
2n,
n)
встречается, например, у пчел: самцы
развиваются из неоплодотворенных
гаплоидных яйцеклеток, самки — из
оплодотворенных диплоидных.
Конкретные механизмы, связывающие
развитие мужского или женского пола с
определенным сочетанием половых хромосом
у разных организмов различен. У человека,
например, пол определяется наличием
Y-xpoмосомы:
в ней есть ТДФ-ген, он кодирует тестикул
- детерминирующий фактор, который
определяет развитие мужского пола.
У дрозофилы же в Y-хромосоме
находится ген фертильности, ответственный
за плодовитость самца, а пол определяется
баллансом числа Х-хромосом и числа
наборов аутосом (это называется половой
индекс)(обычный диплоидный организм
содержит, соответственно, два набора
аутосом). В Х-хромосомах расположены
гены, определяющие развитие по пути
самки, а в аутосомах — по пути самца.
Если отношение количества Х-хромосом
к количеству наборов аутосом равно
0,5(т.е. ХУ: 2 набора аутосом) то развивается
самец, а если — 1(ХХ(2Х): 2 набора аутосом),
то самка.
Помимо нормальных самцов и самок иногда
появляются интерсексы
— особи, по своим половым признакам
занимающие промежуточное положение
между мужским и женским полом (не путать
с гермафродитами!). Это может быть вызвано
как анеуплоидией по половым хромосомам
в гаметах, так и различными нарушениями
(например, гормональными) в процессе
дифференцировки пола.
У большинства
организмов среди потомков получается
50 % самцов и 50 % самок, так как в норме
расщепление по полу подчиняется законам
обычного моногибридного скрещивания
между гомо- и гетерозиготой, для гибридов
которого характерно расщепление в
соотношении 1:1.
Билет№28 Первичные и вторичные половые признаки. Предопределение пола в процессе развития. Нарушение развития пола на примере синдрома Мориса. Наследование, сепленное с полом. Примеры.
Пол характеризуется комплексом признаков, определяемых генами, расположенными в хромосомах. У видов с раздельнополыми особями хромосомный комплекс самцов и самок неодинаков, цитологически они отличаются по одной паре хромосом, ее назвали половыми хромосомами. Одинаковые хромосомы этой пары назвали X(икс)- хромосомами. Непарную, отсутствующую у другого пола- Y (игрек)- хромосомой; остальные, по которым нет различий аутосомами (А). У человека 23 пары хромосом. Из них 22 пары аутосом и 1 пара половых хромосом. Пол с одинаковыми хромосомами XX, образующий один тип гамет (с X- хромосомой), называют гомогаметным, другой пол, с разными хромосомами XY, образующий два типа гамет (с X-хромосомой и с Y-хромосомой), - гетерогаметным. У человека, млекопитающих и других организмов гетерогаметный пол мужской; у птиц, бабочек – женский. X- хромосомы, помимо генов, определяющих женский пол, содержат гены, не имеющие отношения к полу. Признаки, определяемые хромосомами, называются признаками, сцепленными с полом. У человека такими признаками являются дальтонизм (цветная слепота) и гемофилия (несвертываемость крови). Эти аномалии рецессивны, у женщин такие признаки не проявляются, если даже эти гены несет одна из X- хромосом; такая женщина является носительницей и передает их с Х – хромосомой своим сыновьям.
Первичные и вторичные признаки обусловлены генетически, их структура заложена уже в оплодотворенной яйцеклетке задолго до рождения ребёнка. Дальнейшее развитие половых признаков происходит при участии гормонов. К первичным половым признакам относятся те признаки, которые связаны с репродуктивной системой и относятся к строению половых органов. Вторичные половые признаки не участвуют непосредственно в процессе размножения, однако способствуют сексуальному отбору, определяя предпочтения в выборе сексуальных партнёров. Вторичные половые признаки развиваются в период полового созревания.
Цитогенетический метод определения пола. Он основан на микроскопическом изучении хромосом в клетках человека. Применение цито генетического метода позволяет не только изучать нормальную морфологию хромосом и кариотипа в целом, определять генетический пол организма, но, главное, диагностировать различные хромосомные болезни, связанные с изменением числа хромосом или с нарушением их структуры. В качестве экспресс- метода, выявляющего изменение числа половых хромосом, используют метод определения полового хроматина в неделящихся клетках слизистой оболочки щеки. Половой хроматин, или тельце Барра, образуется в клетках женского организма одной из двух Х- хромосом. При увеличении количества Х – хромосом в кариотипе организма в его клетках образуются тельца Барра в количестве на единицу меньше числа хромосом. При уменьшении числа хромосом тельце отсутствует. В мужском кариотипе Y- хромосома может быть обнаружена по более интенсивной люмисценции по сравнению с другими хромосомами при обработке их акрихинипритом и изучении в ультрафиолетовом свете.
Синдро́м нечувстви́тельности к андроге́нам (синдром Морриса) - врожденные эндокринные нарушения полового развития, вызванные мутацией гена, отвечающего за андрогеновый рецептор. Подобные расстройства варьируются в зависимости от структуры и чувствительности аномального рецептора. Большинство форм синдрома включают в себя разный уровень неоконченной маскулинизации и/или бесплодия у XY-индивидов любого гендера.
Билет№29 Нуклеиновые кислоты. Роль ДНК и РНК в реализации наследственной информации в клетке. Доказательства наследственной роли ДНК (опыты Ф.Гриффитса и О.Эвери)
Нуклеи́новая кисло́та (от лат. nucleus — ядро) — высокомолекулярное органическое соединение, биополимер (полинуклеотид), образованный остатками нуклеотидов. Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов и выполняют важнейшие функции по хранению, передаче и реализации наследственной информации. В клетке ДНК содержится главным образом в ядре, а РНК — в цитоплазме. В качестве азотистых оснований в состав нуклеотидов входят пуриновые и пиримидиновые основания (основаниями они названы за свои основные свойства, т.е. способность взаимодействовать с кислотами с образованием солей). ДНК содержит два пурина — а'денин (А) и гуанин (Г) и два пиримидина — цитозин (Ц) и тимин (Т). В состав РНК входят те же самые основания, только вместо тимина — урацил (У). Таким образом, в составе нуклеиновых кислот находятся четыре типа нуклеотидов, различающихся между собой лишь азотистыми основаниями. Причем в ДНК число пуринов всегда равно числу пиримидинов и число А равно числу Т, а число Г числу Ц. Эта особенность связана со структурой молекулы ДНК. Впервые модель молекулы ДНК была предложена в 1953 г. американским ученым-химиком Д. Уотсоном и английским биохимиком Ф. Криком. Согласно этой модели, молекула ДНК состоит их двух спирально закрученных вокруг друг друга нитей.
Способность клеток поддерживать высокую упорядоченность своей организации зависит от генетической информации, которая сохраняется в форме дезоксирибонуклеиновой кислоты (ДНК). Раскрытие роли ДНК в передаче наследственных свойств представляется одним из основных достижений современной биологии. В 1944 г. О. Эвери доказал, что именно ДНК ответственна за изменение (трансформацию) организмов. Это было показано в экспериментах с двумя формами бактерий (пневмококков). Одна из них обладала способностью образовывать капсулу и вызывать заболевание. Вторая форма не образовывала капсулы и не вызывала заболевания. Оказалось, что после проникновения ДНК, выделенной из вирулентных (вызывающих заболевание) клеток, некоторое количество клеток невирулентной формы образовало капсулу, причем эта способность передавалась по наследству. ДНК—это полимер, мономерами которого являются дезоксирибонуклеотиды. В их состав входят углевод дезоксирибоза, фосфорная кислота и азотистые основания четырех типов: два пуриновых — аденин и гуанин и два пиримидиновых — тимин и цитозин. Образование полинуклеотидных цепочек ДНК происходит из трифосфонуклеотидов. Синтез ДНК идет от 5' к 3' концу и катализируется специальными ферментами. Главнейшие из них ДНК-полимеразы, которые последовательно наращивают цепь ДНК, присоединяя к ней дезоксирибонуклеотидные звенья в направлении 5' — к 3'. Именно ДНК-полимеразы на каждом шаге выбирают нужный мономер из четырех, тот, который комплиментарен мономеру материнской цепи ДНК. Однако для начала работы ДНК-полимераз необходима полинуклеотидная цепь рибонуклеиновой кислоты (РНК), называемая затравка. РНК-затравку синтезирует из рибонуклеотидтрифосфатов фермент ДНК-праймаза. В синтезе принимают участие и другие ферменты. ДНК-хеликазы разрывают цепи ДНК, что дает возможность ДНК-полимеразе осуществлять процесс синтеза. ДНК-топоизомеразы раскручивают цепи ДНК и молекулы дестабилизирующего белка, который не позволяет сомкнуться одиночным цепям ДНК. Фермент ДНК-лигаза осуществляет сшивку двух концов цепочки ДНК. Таким образом, в результате совместного действия многих белков осуществляется процесс репликации ДНК, лежащий в основе размножения и развития организма, передачи наследственных свойств. В ДНК заложена информация о структуре белков, свойственных каждому живому организму. Участок ДНК, содержащий всю информацию о программируемом белке, называют ген. Однако в настоящее время установлено, что информационное содержание ДНК значительно богаче. Кроме структурных генов, кодирующих первичную структуру белка, существуют регуляторные участки, которые не кодируют структуру биополимеров, но необходимы для реализации наследственной информации. ДНК содержит информацию и о структуре молекул РНК. Детальная расшифровка структуры ДНК открывает возможность для глубокого проникновения в суть эволюционного процесса. Степень родства организмов может быть установлена с большой точностью путем анализа фрагментов их нуклеиновых кислот. Эти исследования были начаты под руководством академика А.Н. Белозерского
Билет №30 Процесс репликации. Полуконсервативный механизм репликации ДНК. Репликативная вилка. Репликон. Ферменты репликации. Этапы репликации.
Структура ДНК и правило комплементарного спаривания оснований дают представление о возможных механизмах удвоения (репликации) ДНК. Теоретичеки таких механизмов три.
1. Цепи отделяются друг от друга, и каждая служит матрицей для построения комплементарной цепи. В результате синтезируются две молекулы, у каждой из которых одна цепь старая и одна новая. Такой способ репликации ДНК называют полуконсервативным.
2. Если после удвоения одна молекула оказывается состоящей из двух старых цепей, а другая — из двух новых, говорят о консервативном механизме репликации.
3. При дисперсном механизме репликации каждая из двух вновь образованных молекул должна содержать в обеих цепях как новые, так и старые участки. Чтобы узнать, какой из механизмов реализуется в клетках, надо уметь различать старые и новые цепи ДНК. Эту проблему смогли разрешить американские исследователи М Мезельсон и Ф. Сталь в экспериментах по репликации ДН К кишечной палочки. Бактерии в течение большого числа поколений выращивали на среде, содержащей тяжелый изотоп азота 15N. У таких бактерий весь азот в ДНК замещался этим изотопом, и, следовательно, ДНК имела большую плотность, чем ДНК бактерий, выращенных на нормальной среде.
Реплика́ция ДНК — процесс синтеза дочерней молекулыдезоксирибонуклеиновой кислоты на матрице родительской молекулы ДНК. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение. Репликацию ДНК осуществляет сложный ферментный комплекс, состоящий из 15—20 различных белков, называемый реплисомой. Репликация ДНК — ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа: инициация репликации: элонгация: терминация репликации.
Регуляция репликации осуществляется в основном на этапе инициации. Это достаточно легко осуществимо, потому что репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. В геноме таких сайтов может быть как всего один, так и много. С понятием сайта инициации репликации тесно связано понятие репликон. Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномы бактерий, как правило, представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации. Геномы эукариот (а также их отдельные хромосомы) состоят из большого числа самостоятельных репликонов, это значительно сокращает суммарное время репликации отдельной хромосомы. Молекулярные механизмы, которые контролируют количество актов инициации репликации в каждом сайте за один цикл деления клетки, называются контролем копийности. В бактериальных клетках помимо хромосомной ДНК часто содержатся плазмиды, которые представляют собой отдельные репликоны. У плазмид существуют свои механизмы контроля копийности: они могут обеспечивать синтез как всего одной копии плазмиды за клеточный цикл, так и тысяч копий..
Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более распространена двунаправленная репликация. Через некоторое время после начала репликации в электронный микроскоп можно наблюдать репликационный глазок — участок хромосомы, где ДНК уже реплицирована, окруженный более протяженными участками нереплицированной ДНК.
Билет№31 Репарация генетического материала. Дорепликативная репарация – фотореактивация ( световая репарация), темновая эксцизионная репарация. Примеры. Мутация, связанные с нарушением репарации. Мутон. Рекон.
Репарация генетического материала — процесс устранения генетических повреждений и восстановления наследственного аппарата, протекающий в клетках живых организмов под действием специальных ферментов. Способность клеток к репарации генетических повреждений впервые была обнаружена в 1949 году американским генетиком А.Кельнером. В дальнейшем были исследованы многообразные механизмы удаления поврежденных участков наследственного материала, обнаружено, что реперация генетическая присуща всем живым организмам К настоящему времени обнаружено большое количество специализированных репарирующих ферментов, а также гены (см. Ген), контролирующие их синтез в клетках. Доказано, что изменения в этих генах повышают чувствительность организма к неблагоприятным и повреждающим факторам, способствуют возрастанию наследственных изменений — мутаций (см. Мутагенез), возникновению болезней и преждевременному старению. Детально изучены две формы репапрации генетической — фотореактивация и темновая репарация.Фотореактивация, или световое восстановление, была обнаружена в 1949 г. А. Кельнер, изучая биологическое действие радиации в экспериментах на микроскопичских грибах и бактериях, обнаружил, что клетки, подвергшиеся одинаковой дозе ультрафиолетового облучения, выживают значительно лучше, если после облучения в темноте их поместить в условия обычного естественного освещения. Исходя из этого, было высказано предположение, что на свету происходит устранение части поврелсдений генетических структур клеток, возникающих под действием ультрафиолетового облучения.
Оказалось, что ультрафиолетовое облучение обладает способностью нарушать структуру молекул дезоксирибонуклеиновой кислотыты (сокращенно ДНК — см. Нуклеиновые кислоты), несущих генетическую информацию. Молекула ДНК содержит четыре типа так называемых азотистых оснований: аденин, гуанин, цитозин и тимин — и состоит из двух нитей, закрученных в спираль. Нередко в одной нити одинаковые основания располагаются рядом. Под действием ультрафиолетового облучения в части азотистых оснований разрываются химические. Для «лечения», устранения этих повреждений в некоторых клетках имеются специальные ферменты, названные фотореактивирующими. Однако «лечебный эффект» фотореактивирующих ферментов — расщепление сцепленных участков молекулы ДНК и восстановление ее исходной нормальной структуры — проявляется только при участии световой энергии. Тогда отсюдова, свет играет в этих процессах роль активирующего фактора, запускающего реакцию фотореактивации. До сих пор это остается единственным примером биохимических реакций, в которых активатором выступает световая энергия.
Темновая репарация, в отличие от фотореактивации, универсальна. Она устраняет различные структурные повреждения ДНК, появляющиеся в результате разнообразных радиационных и химических воздействий. Способность к темновой репарации обнаружена у всех клеточных систем и организмов. Оказалось, что механизмы темновой репарации принципиально отличны от механизма фотореактивации. Первое отличие заключается в том, что если во время реакции на свету фотореактивирующий фермент расщепляет сцепленные ультрафиолетовым облучением участки молекулы ДНК, то в ходе темновой репарации поврежденные участки удаляются из молекулы ДНК. Второе отличие связано с числом «вылечиваемых» повреждений. Фотореактивирующий фермент активен в отношении только одного типа повреждений ДНК — образования димеров тимина под действием ультрафиолетового облучения. Ферменты же, осуществляющие темновую репарацию, способны устранять различные структурные нарушения ДНК, появляющиеся вследствие всевозможных воздействий на клетки — и химических, и радиационных. В результате темновой репарации осуществляется своеобразное молекулярное «хирургическое» вмешательство: поврежденные участки «вырезаются», а образовавшиеся «бреши» заполняются путем локального (местного) синтеза или обмена участками между поврежденной и неповрежденной нитями ДНК, в результате чего и восстанавливается ее исходная нормальная структура. Темновая репарация осуществляется под контролем большого числа ферментов, каждый из которых отвечает за определенный этап этого сложного процесса. Детально изучены два типа темновой репарации — эксцизионная и пострепликативная. При эксцизионной репарации поврежденный участок ДНК вырезается и замещается до начала очередного цикла размножения клетки, точнее до начала удвоения (репликации) молекул ДНК. Биологический смысл этого процесса состоит в том, чтобы предупредить закрепление у потомства наследственных изменений (мутаций) и последующее размножение измененных форм. Эксцизионная репарация — наиболее экономичная и эффективная форма репарации генетической. установлено, что при ее нормальном функционировании у микроорганизмов до начала репликации ДНК удаляется до 90% имеющихся генетических повреждений, из клеток высших организмов — до 70%. Эксцизионная репарация осуществляется в несколько этапов.Сначала специальный фермент «надрезает» одну из нитей ДНК, вблизи от поврежденного участка, затем поврежденный участок удаляется полностью, а образовавшуюся «брешь» заполняют специальные ферменты (ДНК-поли-меразы), которые поставляют недостающие звенья, заимствуя их из неповрежденной нити. Способность к эксцизионной репарации установлена у клеток микроорганизмов, высших растений и животных, а также у человека.
Связь мутаций с репарацией ДНК Мутации возникают лишь тогда, когда репарационный механизм по каким-то причинам не работает или не справляется с устранением повреждений. Мутации, возникающие в генах, кодирующих белки, ответственные за репарацию, могут приводить к многократному повышению (мутаторный эффект) или понижению (антимутаторный эффект) частоты мутирования других генов. Так, мутации генов многих ферментов системы эксцизионной репарации приводят к резкому повышению частоты соматических мутаций у человека, а это, в свою очередь, приводит к развитию пигментной ксеродермы и злокачественных опухолей покровов.
В домолекулярной генетике для обозначения наименьшего элемента в структуре ДНК, который может быть подвержен спонтанной или индуцированноймутации, применялся особый термин рекон. В настоящее время показано, что таким наименьшим элементом является один нуклеотид (или одно азотистое основание в составе нуклеотида), поэтому данный термин более не употребляется. Для определения понятия единица мутации применялся термин мутон. В настоящее время показано, что фенотипически мутация может проявиться даже при замене одного нуклеотида (или азотистого основания в составе нуклеотида), таким образом, термин мутон соответствует одному нуклеотиду.
Билет№32 Репарация генетического материала. SOS – система. Пострепликативная репарация.
Репарация генетического материала — процесс устранения генетических повреждений и восстановления наследственного аппарата, протекающий в клетках живых организмов под действием специальных ферментов. Способность клеток к репарации генетических повреждений впервые была обнаружена в 1949 году американским генетиком А.Кельнером. В дальнейшем были исследованы многообразные механизмы удаления поврежденных участков наследственного материала, обнаружено, что реперация генетическая присуща всем живым организмам К настоящему времени обнаружено большое количество специализированных репарирующих ферментов, а также гены (см. Ген), контролирующие их синтез в клетках. Доказано, что изменения в этих генах повышают чувствительность организма к неблагоприятным и повреждающим факторам, способствуют возрастанию наследственных изменений — мутаций (см. Мутагенез), возникновению болезней и преждевременному старению. Детально изучены две формы репапрации генетической — фотореактивация и темновая репарация.
При прохождении репликативного комплекса через некодирующий или ошибочно кодирующий поврежденный участок ДНК наблюдается включение в синтезируемую цепь случайных или соответствующих мутантному участку нуклеотидов. Затем репликация ДНК продолжается в обычном режиме. Таким образом, появляются поврежденные участки ДНК.
У E. coli в ответ на повреждения ДНК происходит координированная экспрессия большого числа генов (прмерно 26 белков). Такая реакция бактериальных клеток на генотоксические воздействия получила название SOS-ответа , а процесс образования мутаций - SOS- мутагенеза. В индукции SOS-ответа у E. coli определяющую роль играют ген lexA и ген recA .Белок LexA является репрессором гена recA и более 20 других генов и оперонов, составляющихSOS-регулон . В ответ на повреждение ДНК или ингибирование репликации, при прохождении ДНК- полимеразой поврежденного участка ДНК вырабатывается SOS-сигнал. Итак, SOS-система - генетическая программа изменения ДНК путем ее мутагенной репарации или рекомбинации с целью адаптации клетки к изменившимся внешним условиям, результатом чего является наращивание ее эволюционного потенциала. Пострепликативная репарация — последняя возможность для клетки устранить имеющиеся генетические повреждения, защитить потомство от изменения наследственных признаков. Если в ДНК возникает так много повреждений, что в ходе эксцизионной репарации клетка не успевает их полностью устранить, или если повреждены гены, определяющие возможность эксцизионной репарации, то в процессе размножения (удвоения, репликации) ДНК в дочерних нитях на месте повреждений, имеющихся в материнской нити,образуются «бреши». Это происходит в силу того, что фермент, ведущий репликацию ДНК (синтез дочерней нити на материнской нити ДНК), не может «прочесть» искаженную информацию в поврежденной точке материнской нити. Поэтому, доходя до поврежденного места, оставшегося неисправленным во время эксцизионной репарации, этот фермент останавливается, затем медленно (со скоростью в сотни раз меньшей, чем обычно) проходит через зону повреждения и возобновляет нормальный синтез дочерней нити, отступя от этого места. Так происходит во всех точках, где материнская нить ДНК остается поврежденной к началу репликации. Конечно, если число повреждений слишком велико, репликация останавливается полностью и клетка погибает. Но и существовать с молекулами ДНК, несущими бреши, клетка долго не может. Поэтому после репликации, но перед делением клетки начинается процесс пострепликативной репарации. Перед делением клетки в ней образуются две двунитевые молекулы ДНК. Если одна из них несет в какой-либо точке повреждение в одной нити и брешь в противоположной нити, то в другой двунитевой молекуле ДНК обе нити в данной точке будут нормальными. В этом случае может произойти обмен участками ДНК — рекомбинация (см. Ген, обмен генами): неповрежденный участок будет вырезан из нормальной молекулы ДНК и вставлен на место поврежденного участка в другой молекуле, благодаря чему поврежденный генетический материал будет заменен нормальным. Вслед за этим спец. ферменты (ДНК-полимеразы) заделают «бреши» (теперь они смогут это сделать, т. к. в обеих молекулах в данном месте повреждения будут отсутствовать), вновь синтезированные и старые нити будут соединены друг с другом, и исходная структура ДНК будет в результате этого полностью восстановлена. В соответствии с природой процесса, связанного с осуществлением рекомбинации, этот тип пострепликативной репарации называют также рекомбинационным.
Билет№33 Характеристика наследственного материала в митотическом цикле клетки. Химический состав и структурная организация хроматина. Морфология хромосом. Хромосомные типы “ламповых счеток”. Политенные хромосомы.
Клеточный цикл — это период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти.
Важным компонентом клеточного цикла является митотический (пролиферативный) цикл —комплекс взаимосвязанных и согласованных во времени событий, происходящих в процессе подготовки клетки к делению и на протяжении самого деления. Биологическое значение митотического цикла состоит в том, что он обеспечивает преемственность хромосом в ряду клеточных поколений, образование клеток, равноценных по объему и содержанию наследственной информации. Таким образом, цикл является всеобщим механизмом воспроизведения клеточной организации эукариотического типа в индивидуальном развитии. Главные события митотического цикла заключаются в редупликации (самоудвоении) наследственного материала материнской клетки и в равномерном распределении этого материала между дочерними клетками. Значат для митотического цикла наиболее важен S период. Хроматин имеет вид глыбок, гранул и нитей. Химический состав хроматина: 1) ДНК (30–45%), 2) гистоновые белки (30–50%), 3) негистоновые белки (4–33%), следовательно, хроматин является дезоксирибонуклеопротеидным комплексом (ДНП). В зависимости от функционального состояния хроматина различают: гетерохроматин (5) и эухроматин (6). Эухроматин — генетически активные, гетерохроматин — генетически неактивные участки хроматина. Эухроматин при световой микроскопии не различим, слабо окрашивается и представляет собой деконденсированные (деспирализованные, раскрученные) участки хроматина. Гетерохроматин под световым микроскопом имеет вид глыбок или гранул, интенсивно окрашивается и представляет собой конденсированные (спирализованные, уплотненные) участки хроматина. Хроматин — форма существования генетического материала в интерфазных клетках. Во время деления клетки (митоз, мейоз) хроматин преобразуется в хромосомы.Хромосомы типа ламповых щеток, это специальная форма хромосом, которую они приобретают в растущих ооцитах (женских половых клетках) большинства животных, за исключением млекопитающих. В растущих ооцитах всех животных, за исключением млекопитающих, во время протяженной стадии диплотены профазы мейоза I активная транскрипция многих последовательностей ДНК приводит к преобразованию хромосом в хромосомы, по форме напоминающие щетки для чистки стёкол.
Политенные хромосомы — гигантские скопления объединённых хроматид, возникающие в некоторых типах специализированных клеток. Репликация ДНК в этих клетках не сопровождается делением клетки, что приводит к накоплению вновь построенных нитейДНК. Как клетки, так и политенные хромосомы значительно увеличиваются в размерах, что облегчает наблюдение и позволяло изучать активность генов
Билет№34 Кариотип и идеограмма хромосом человека. Строение, типы хромосом. Характеристика диплоедного и гаплоидного набора хромосом. Методы анализа фотокариограммы. Группы хромосом в кариотипе человека.
Кариотип – диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида, являющийся видоспецефическим признаком и характеризующийся определённым числом и строением хромосом. Если число хромосом в гаплоидном наборе половых клеток обозначить п, то общая формула кариотипа будет выглядеть как 2п, где число п различно для разных видов.