

1. Отступать от правил поведения, описанных выше;
2. Ходить в грязной одежде и обуви;
3. Действовать самовольно и наугад.
Лекция 4. Типичные ошибки при проведении ПЦР
Метод ПЦР состоит из большого числа последовательных и аккуратных действий. Способность контролировать каждый свой шаг в этом методе приходит со знаниями и опытом.
Очень важным моментом является воспроизводимость результатов ПЦР. Для того чтобы этот показатель был высоким, нужно стандартизовать условия проведения ПЦР. Условия должны быть одинаковыми для всех проб.
Рассмотрим типичные источники ошибок, встречающиеся на каждом из этапов ПЦР.
Этап выделения ДНК.
1. Неправильное хранение биологического материала.
2. Загрязнение проб.
3. Нарушение очередности действий в этапе выделения ДНК.
4. Истечение срока годности реактивов для выделения ДНК.
Этап амплификации ДНК.
1. Нарушение пропорций при смешивании реактивов для амплификации.
2. Использование праймера, не способного отжигаться на ДНК данного организма.
3. Загрязнение проб ДНК из соседних пробирок (при отсутствии смены наконечников).
4. Истечение срока годности реактивов для амплификации ДНК.
Этап детекции продуктов амплификации.
1. Использование силы тока и напряжения, неподходящих для электрофореза ДНК.
2. Неправильное подключение камеры для электрофореза (ДНК-фрагменты всегда двигаются к аноду).
3. Испорченный трис-боратный буфер. Это возможно при неправильном хранении либо при продолжительном использовании буфера.
4. Неправильное приготовление и старение агарозного геля (высокая хрупкость, сомкнутые стенки лунок и т. д.).
Лекция 5. Применение ПЦР для разных видов научно-исследовательской работы
Использование ПЦР-метода обычно происходит в следующих двух типах ситуаций.
1. Используется специфические праймеры, которые будут отжигаться на определенном участке ДНК, заранее известном исследователю (фото 4). То есть, известны места отжига праймера и расстояния между сайтами отжига праймеров. Это может быть определенный ген, теломеры хромосом, палиндромы и т. д. Для того, чтобы знать последовательность определенного участка ДНК, нужно прежде провести секвенирование данного фрагмента. Когда последовательность установлена и подобран праймер, можно проводить ПЦР.

M F
М – маркер молекулярных весов ДНК от 200 до 1000 п.о. с шагом в 100 п.н., F – фрагмент вирусной ДНК длиной 317 п.н.
Фото 4. Продукт амплификации вирусной ДНК со специфическими праймерами, выделенной из полиэдров вируса ядерного полиэдроза непарного шелкопряда
Специфические праймеры используется, например, в диагностике инфекционных заболеваний. Для этого подбирается (например, в GeneBank) и синтезируется праймер, который будет отжигаться только на ДНК определенного возбудителя заболевания.
2. Используется праймер (праймеры), который будет отжигаться случайно на ДНК. Такая разновидность ПЦР называется RAPD-PCR (Random Amplified Polymorphic DNA, случайно амплифицированная ДНК). Часто используется для изучения генетического полиморфизма в популяциях. При этом невозможно предсказать точно ни длину наработанных фрагментов, ни их число (фото 5). Кроме этого, всегда существует вероятность, что используемый праймер не сработает. Поэтому перед началом исследований подбирают подходящий праймер.

1.1, 2.1,3.1, 4.1, 5.1, 6.1, 7.1 – исследовалась поверхность яиц на наличие вирусной ДНК; 1.2, 2.2, 3.2, 4.2, 5.2, 6.2, 7.2 – исследовалось содержимое яиц на наличие вирусной ДНК; М – маркер молекулярных весов ДНК от 100 до 1000 п.о. с шагом в 100 п.о.; 00, 01, 02, 03 – условные зоны продуктов амплификации вирусной ДНК
Фото 5. Продукты амплификации ДНК вируса ядерного полиэдроза с поверхности и изнутри яиц непарного шелкопряда (праймер OPA-08)
Для примера смоделируем исследовательскую ситуацию. Например, существует организм А, на который действует фактор B (биотический или абиотический). Если произведено секвенирование генома организма А, найдены гены, отвечающие за взаимодействие с фактором B, подобраны праймеры к ним (генам), то лучше всего проводить исследование типа 1. Если отсутствует информация о геноме организма А (или ее очень мало), то проводиться исследование типа 2.
Далее (после ПЦР), например, в случае проведения исследования второго типа, в зависимости от полученных фрагментов ДНК, ведется поиск маркеров взаимодействия между организмом А и фактором B. Например, может оказаться, что наличие маркера длиной Х усиливает влияние фактора В на организм А, а наличие маркера длиной Y, наоборот, уменьшает это влияние.
Теоретически можно рассчитать максимальное количество ампликонов (A) разной длины, которые будут амплифицироваться (второй тип исследования). При этом надо знать длину исследуемой ДНК (D) и длину праймера (L).
Для большинства существующих праймеров минимальный возможный размер ампликона будет равен 2L. Математически это можно представить в следующем виде:
D = A(A – 1)/2 + 2LA + M,
где М – остаток ДНК, на которой отжиг праймера не способен привести к амплификации фрагмента ДНК с длиной A+1;
M < 2L + A.
Вероятность такого события (образования максимального количества ампликонов разной длины с учетом 4 различных гетероциклических оснований) может быть описана математически:
P = 4-2LA.
Из этой формулы следует, что чем больше длина праймера, тем меньше вероятность амплификации фрагментов разной длины.
Лекция 6. Подбор праймеров
Концентрация праймеров обычно варьирует в пределах 0,1-1 мкМ. Более высокая концентрация праймеров может приводить к образованию неспецифических продуктов.
Праймеры подбирают с учетом следующих требований:
а) содержание оснований Г и Ц в праймере должно находиться в пределах 40-60 %;
б) основания Г и Ц должны быть равномерно распределены по всей длине праймера;
в) желательно чтобы на 3’-конце праймера находилось основание Г или Ц, так как это увеличивает эффективность ПЦР;
г) следует избегать наличия 3 и более оснований Г или Ц на 3’-конце праймера, поскольку это может приводить к ошибочному спариванию праймера с ГЦ-богатыми участками ДНК;
д) праймеры не должны формировать стабильных дуплексов сами на себя или друг с другом;
е) участки отжига должны быть уникальными, т.е. праймеры не должны иметь длинных комплементарных участков вне специфических мест отжига;
ж) температура плавления (Тm) данной пары праймеров должна быть сходной и находиться в пределах 55-65 оС.
Для ориентировочного подсчета температуры плавления используют формулу:
Тm = 2(А+Т) + 4(Г+Ц),
где А, Т, Г, Ц, – количество соответствующих нуклеотидов в праймере. Более точные результаты можно получить при использовании формулы:
Тm = 81,5 + 16,6(lgM) + 0,41(%Г+Ц)–(600/L),
где М – ионная сила раствора (в молях/л), L – длина олигонуклеотида. Эта формула пригодна для олигонуклеотидов длиной 14-70 оснований (Сулимова и др., 2006).
Лекция 7. «Золотая середина» концентраций веществ в ПЦР-смеси
Taq-полимераза
Является термофильной ДНК-полимеразой (из Thermis aquaticus).
Точность Taq-полимеразы зависит от концентрации Mg2+, dNTP, сбалансированности dNTP, pH. В среднем, ошибка чуть ниже, чем 1 замена на 100-300 п.н.
Количество ДНК-полимеразы должно быть оптимально. Если используется слишком маленькое количество Taq-полимеразы, выход продукта снижается, причем это снижение тем серьезнее, чем больше размер продукта. При 2-4-кратном избытке Taq-полимеразы в начале реакции появляются высоко- и низкомолекулярные продукты, а при 4-16-кратном превышении продукт становится низкомолекулярным. Необходимо иметь в виду, что Taq-полимераза обладает 5’-3’ экзонуклеазной активностью. Эта активность может вызвать проблемы, обусловленные деградацией 5’ концов продукта.
Дезоксинуклеотидтрифосфаты (dNTP)
Диапазон исследуемых концентраций dNTP – 0,05-0,5 mM. Чаще всего концентрация dNTP в реакционной смеси составляет 0,2 mM. Увеличение концентрации dNTP выше 0,2 mM приводит к снижению специфичности и точности ПЦР. Снижение концентрации dNTP снижает частоту ошибок фермента, однако приводит к снижению выхода ПЦР-продукта.
Ионы магния (Mg2+)
Mg2+ образует комплексы с dNTP (именно эти комплексы являются субстратом для Taq-полимеразы), праймерами и матрицей ДНК и поэтому концентрация Mg2+ влияет на отжиг праймеров, температуру диссоциации матричной и амплифицированной ДНК , специфичность продукта, активность и точность ДНК-полимеразы. Для нормальной работы ДНК-полимеразы необходимо наличие свободных ионов магния, а не только тех, которые связаны с ДНК-матрицей, праймерами и dNTP. Поэтому концентрация Mg2+ в реакционной смеси должна быть на 0,5-2,5 mM выше общей концентрации dNTP. При концентрации dNTP в 0,2 mM концентрация Mg2+ обычно варьирует в пределах 1,25-1,75 mM. Слишком низкая концентрация Mg2+ - низкий выход ПЦР-продукта, слишком высокая – продукты неспецифической амплификации.
Матрица для ПЦР
Количество ДНК-полимеразы в реакционной смеси может меняться в широком интервале: от 0,01 до 1 мкг на 100 мкл реакционной смеси – в зависимости от типа матрицы. Для геномной ДНК млекопитающих это количество составляет 0,5-1 мкг, для геномной ДНК бактерий – 50 нг. Необходимо иметь в виду, что при низкой концентрации матрицы необходимо использовать силиконизированные пробирки для предотвращения неспецифической абсорбции ДНК на стенках пробирки.
В определенных условиях можно получить амплификацию с единичной копии матрицы. При повышении концентрации ДНК более 1 мкг/100 мкл происходит уменьшение выхода специфического продукта ПЦР из-за образования неспецифических продуктов.
Для обеспечения максимального выхода и специфичности продукта в каждом конкретном случае использования ПЦР необходимо оптимизировать условия реакции (подобрать концентрацию праймеров, ионов Mg2+, дезозоксинуклеотидов, матричной ДНК и полимеразы, подобрать время инкубации и температуру, состав инкубационной смеси (Сулимова и др., 2006).
Занятие 1. Выделение ДНК
Для этапа выделения ДНК используется, как правило, свежий биологический материал, в котором ДНК находится в целостном состоянии. Все факторы, которые приводят к разрыву ковалентных связей в исследуемой ДНК и загрязняют пробы чужеродной ДНК, обусловливают ошибки при проведении ПЦР и должны быть упразднены. К таковым, прежде всего, нужно отнести высокую температуру (40–50 оC и выше), электромагнитное излучение (ультрафиолет, рентгеновские лучи), радиационное излучение (α, β, γ-лучи), присутствие жизни на исследуемом материале (бактерии, грибы). Наиболее оптимальным местом хранения ДНК является морозильная камера. При температуре -5 – -20 оС ДНК в клетках может сохраняться достаточно длительное время, не разрушаясь. Для хранения материала в течение 1-2 суток используется холодильник (2-8 оC).
Для выделения ДНК лучше всего подходит ткань, где клетки активно делятся и растут.
Мертвая ткань и ткани опорно-трофической функции, где ДНК встречается в полуразрушенном состоянии, дают слабый выход ампликонов (фото 6).
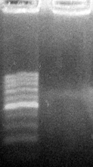
M О
М – маркер молекулярных весов ДНК (длина фрагментов от 100 до 1000 п. н. с шагом в 100 п. н.)
О – отсутствие продуктов амплификации ДНК
Фото 6. Отрицательная ПЦР-реакция (праймер OPA 14) с ДНК, выделенной из плавников пеленгаса
Объем материала, необходимый для выделения достаточного количества ДНК, зависит от размера клеток. Чем меньше размер клеток, тем меньше необходимо исследуемого вещества. Если ПЦР состоит из 40 циклов, то теоретически достаточно и одной клетки для того, чтобы обнаружить наработанные ДНК-фрагменты. Чем меньше исследуемого материала (количества клеток), тем аккуратней нужно проводить исследование, чтобы не загрязнить пробу.
При выделении ДНК используются стандартные наборы реактивов. Например, комплект «ДНК-сорб-А» фирмы «АмплиСенс» (Москва, Россия) для выделения ДНК, который имеет следующий состав:
1. лизирующий раствор;
2. отмывочный раствор;
3. сорбент универсальный;
4. ТЕ-буфер для элюции ДНК.
Хранится набор при температуре 2-25 оС. Рассчитан на выделение ДНК из 100 проб.
Порядок работы.
1. Отобрать необходимое количество одноразовых пробирок. Промаркировать пробирки.
2. Растереть пробы в ступке с добавлением 100 мкл дистиллята и перенести пробы по отдельным пробиркам.
3. Лизирующий раствор (если он хранился при 2-8 оС) прогреть при 65 оС до полного растворения кристаллов. В пробирки добавить по 300 мкл лизирующего раствора.
4. Пробы тщательно перемешать на вортексе и прогреть 5 мин при 65 оС. Процентрифугировать 5 с при 5 тыс. об/мин на центрифуге. Если проба растворилась не полностью, процентрифугировать пробирку на микроцентрифуге 5 мин при 12 тыс. об/мин и использовать для выделения ДНК надосадочную жидкость, перенеся ее в новую пробирку.
5. Тщательно ресуспендировать сорбент на вортексе. В каждую пробирку отдельным наконечником добавить по 20 мкл ресуспендированного сорбента. Перемешать на вортексе, поставить в штатив (планшет) на 2 минуты, еще раз перемешать и оставить в штативе на 5 минут.
6. Осадить сорбент в пробирках центрифугированием при 5 тыс. об/мин в течение 30 с. Удалить супернатант, используя дозатор с переменным объемом и меняя наконечник для каждой пробирки.
7. Добавить в пробирки по 500 мкл раствора для отмывки, перемешать на вортексе до полного ресуспендирования сорбента, процентрифугировать 30 с при 10 тыс. об/мин на микроцентрифуге.
8. Удалить супернатант, используя дозатор с переменным объемом и меняя наконечник для каждой пробирки.
9. Повторить отмывку еще раз, следуя пункту 8., удалить супернатант полностью.
10. Поместить пробирки в термостат при 65 оС на 5-10 мин для подсушивания сорбента (осадок приобретает цвет мела). При этом крышки пробирок должны быть открыты.
11. В пробирки добавить по 100 мкл ТЕ-буфера для элюции ДНК. Перемешать на вортексе. Переместить в термостат при 65 оС на 5 мин, периодически встряхивая на вортексе.
12. Процентрифугировать пробирки при 12 тыс. об/мин в течение 1 мин на микроцентрифуге. Супернатант содержит очищенную ДНК, пробы готовы к постановке ПЦР.
Очищенную ДНК можно хранить в течение недели при 2-8 оС и в течение 1 года при минус 20 оС. Эффективность выделения ДНК составляет 50-70 %.
Задание 1.1.
Выделить ДНК из листьев комнатных растений.
Задание 1.2.
Выделить ДНК из полиэдров вируса ядерного полиэдроза непарного шелкопряда.
Занятие 2. Амплификация ДНК
Основой данного этапа ПЦР является программируемый амплификатор. Амплификатор в зависимости от стадии изменяет температурный режим пробирок и временной интервал.
Амплификация ДНК включает в себя 3 стадии*:
1. денатурация ДНК (0,5-1 минута, 93 оС),
2. отжиг праймеров (0,5-1 минута, 35-40 оС),
3. наращивание цепи (1 минута, 72 оС).
*Температура и продолжительность цикла подходят для праймеров типа OPA.
Амплификация фрагмента определенной длины, который образуется при классическом отжиге праймера, изображенном на рис. 2, происходит по следующей математической формуле:
F= 2(n-1) + 22(n-2) + 23(n-3) + 24(n-4) + 25(n-5) +... +2n-1 (a) ,
где n – количество циклов, F – количество одноцепочечных фрагментов в начале цикла n+1 (денатурация).
Количество нарабатываемых двухцепочечных фрагментов описывается следующей математической формулой:
F= 2 1(n-2) + 22(n-3) + 23(n-4) + 24(n-5) + 25(n-6) +... +2n-2 (b),
где n- количество циклов, F-количество двухцепочечных фрагментов.
Рассмотрим значения формулы b для каждого цикла:
1-й цикл – –
2-й цикл – 0
3-й цикл – 2n-4
4-й цикл – 6n-16
5-й цикл – 14n-48 и т. д.
Каждое значение фрагментов для любого цикла можно представить в следующем виде:
(2n-1-2)n – (2n-1-2)(n-2) – 2(n-2) = F,
2(2n-1-2) – 2(n-2) = F,
2(2n-1 - n) = F.
Количество ампликонов с каждым циклом(n) увеличивается почти в два раза. При n→ ∞ это можно записать в следующем виде:
lim 2n-1 – 2(n – 1)/ 2n – 2n = 1/2
n→ ∞
Для проведения данного этапа ПЦР (амплификации) используют разные приемы. Рассмотрим два из них.
«Горячий старт» («hot start») – прием, при котором пробирки с реагентами помещаются в амплификатор, нагретый до температуры 93-94 оС. Денатурация ДНК 1-го цикла должна протекать 2-2,5 минуты. Остальные циклы протекают согласно описанным выше температурным и временным константам. Данный прием позволяет избежать неспецифического отжига праймеров. Существуют разные модификации данного приема (например, с использованием воска).
Обычный прием предусматривает помещение пробирок в амплификатор при комнатной температуре. После достижения амплификатором температуры в 94 оС начинается процесс денатурации и т. д.
Для проведения этапа амплификации ДНК используются стандартные наборы реактивов. Примером может служить набор «АмплиСенс – 200 -1» фирмы «АмплиСенс» (Москва, Россия). В его состав входят:
1. смесь нуклеотидтрифосфатов, 2 mM;
2. 5-кратный реакционный буфер, без Mg2+;
3. сульфат магния, 50 mM;
4. деионизованная вода, стерильная, автоклавированная;
5. Taq-полимераза, 5 ед/мкл;
6. воск для ПЦР, 14%-ный, плавление при 37 оС;
7. минеральное масло, вазелиновое;
8. исследуемая ДНК.
Такой набор рассчитан на 200 реакций в объеме 25 мкл. Хранится набор при температуре 2-8 оС.
Смешивание реактивов проводится в такой пропорции (возможны вариации):
1. смесь нуклеотидтрифосфатов – 2,5 мкл;
2. 5-кратный реакционный буфер – 5 мкл;
3. сульфат магния – 1,5 мкл;
4. деионизованная вода – 3 мкл;
5. Taq-полимераза – 0,5 мкл;
6. минеральное масло – 11,5 мкл;
7. исследуемая выделенная ДНК – 5мкл;
8. праймер – 1 мкл.
Общий объем: 30 мкл.
Амплификация ДНК длится приблизительно 2,5 – 3,5 часа (30-40 циклов).
Количество праймера, необходимое для амплификации одного ДНК-фрагмента с одной молекулы ДНК в течение n циклов (рис. 2), описывается математической формулой:
P=2n+1 – 2, где n - количество циклов, а Р - количество молекул праймера.
Порядок работы.
1. Промаркировать пробирки.
2. Смешать реактивы, исследуемую ДНК и праймер в пропорции описанной выше. Процентрифугировать на вортексе в течение 1 минуты, чтобы на стенках пробирок не осталось реагентов.
3. Выбрать подходящую программу на амплификаторе, загрузить пробирки в амплификатор и запустить амплификацию.
4. По окончанию амплификации можно приступать к 3-му этапу ПЦР (детекции продуктов амплификации).
Пробирки с наработанными фрагментами ДНК можно хранить при температуре 2-8 оС в течение двух недель.
Задание 2.1.
Провести амплификацию ДНК, выделенной из листьев комнатных растений с использованием праймеров серии OPA.
Задание 2.2.
Провести амплификацию ДНК, выделенной из полиэдров вируса ядерного полиэдроза непарного шелкопряда со специфическими праймерами: а) 5’- GCC GGC GGA ACT GGC CCA -3’, б) 5’- CGA CGT GGT GGC ACG GCG -3’.
Занятие 3. Детекция продуктов амплификации
Является финальной стадией ПЦР. Для детекции амплифицированных фрагментов ДНК используется метод горизонтального электрофореза в агарозном геле с использованием трис-боратного буфера. Для того, чтобы визуально обнаружить ДНК-фрагменты в трис-боратный буфер добавляют бромистый этидий*. Это вещество способно встраиваться в двухцепочечные молекулы ДНК и флуоресцировать под ультрафиолетовыми лучами**. Именно флуоресцирующие ДНК-фрагменты являются целью исследователя, применяющего ПЦР-метод (фото 7). При помощи цифрового фотоаппарата ДНК-фрагменты записываются и могут быть проанализированы.
*Бромистый этидий – сильный канцероген, способный проникать через кожу в организм (рис. 4). Следует уделять особое внимание при работе с ним. Для работы с бромистым этидием используются перчатки.
|

Рис. 4. Химическое строение бромистого этидия
**Ультрафиолетовые лучи опасны для глаз. Поэтому при работе с ультрафиолетовой лампой нужно избегать прямого попадания лучей в глаза и использовать защищающие глаза специальные очки или маски. Материалом для очков и масок может служить стекло с примесями окиси железа(Fe2O3), например, оконное стекло (0. 1 % Fe2O3).
Для этапа детекции продуктов амплификации используются следующие химические вещества:
1. агароза;
2. трис-боратный буфер*;
3. бромистый этидий;
4. маркер молекулярных весов.
*Трис-боратный буфер можно приготовить самостоятельно.
Необходимы следующие химические вещества:
Трис-(оксиметил)-аминометан – 10.78 г,
Борная кислота – 5.503 г,
EDTA, pH 8.0 – 4.384 мл,
дистиллят – 86.3 мл,
бромистый этидий – 10-15 мкл (10 мг/мл).
Смешать химические вещества и размешать до полного растворения. Довести дистиллятом объем раствора до литра. Хранить трис-боратный буфер нужно при температуре 18-25 оС.
Порядок работы.
1. Приготовить 1.8%-ный агарозный гель на основе трис-боратного буфера. Поместить его в камеру для электрофореза.
2. Залить гель трис-боратным буфером, содержащим бромистый этидий.
3. Нанести по 5 мкл каждой пробы в отдельные лунки. Нанести в отдельную лунку маркер.
4. Подключить камеру для электрофореза к источнику электрического питания (50 mA, 100 V).
Через 30-70 минут после начала электрофореза достать гель и сфотографировать его под ультрафиолетовой лампой, используя специальные очки для защиты глаз (фото 7).
Количество наработанных фрагментов ДНК определяют качество визуальной детекции ампликонов. Сколько же нужно ампликонов одного фрагмента ДНК для визуальной детекции? Для фрагмента ДНК длиной от 100 до 200 п. н. достаточно 108–109 двухцепочечных ампликонов. Теоретически, это приблизительно 30 циклов ПЦР на ДНК, выделенной из одной клетки.

1 2 3 М
М-маркер (длина фрагментов от 100 до 1000 п. н.; шаг– 100 п. н.),
1, 2, 3 – фрагменты ДНК дуба пушистого.
Фото 7. Продукты амплификации ДНК дуба пушистого (OPA 14)
Чем меньше длина двухцепочечного ДНК-фрагмента, тем слабее он флуоресцирует. Это объясняется тем, что в короткий ДНК-фрагмент встраивается меньшее количество молекул бромистого этидия, чем в длинный.
Бромистый этидий обладает интересной особенностью, за которую его и начали использовать в ПЦР-методе. При встраивании бромистого этидия в двухцепочечную молекулу ДНК, его способность к флуоресценции возрастает в 20 раз. Максимум спектра поглощения бромистого этидия находится в области ультрафиолетовых лучей.
Статистическую обработку данных ПЦР-анализа можно проводить при помощи различных компьютерных программ:
Treecon for Windows (version 1.3 b), MAPMAKER 3.0, Граф-анализ и др.
Задание 3.1.
Провести детекцию продуктов амплификации ДНК комнатных растений (праймеры серии OPА).
Задание 3.2.
Провести детекцию продуктов амплификации ДНК вируса ядерного полиэдроза непарного шелкопряда (специфические праймеры).