

Основные положения хромосомной теории наследственности
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:
Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
Гены расположены в хромосоме в линейной последовательности.
Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
Каждый биологический вид характеризуется определенным набором хромосом — кариотипом.
Сцепленное наследование
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы до эры молекулярной биологии было изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. То, что число генов у высших организмов составляет несколько тысяч, было ясно уже У. Сэттону в начале XX века. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе.
Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием. Число групп сцепления соответствует гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. (У особей гетерогаметного пола, например, у самцов млекопитающих, групп сцепления на самом деле на одну больше, так как X- и У-хромосомы содержат разные гены и представляют собой две разные группы сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин — 24).
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и ab) в равных количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.
Было установлено, однако, что кроме обычных (некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми комбинациями генов — Ab и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной возникновения таких гамет является обмен участками гомологичных хромосом, или кроссинговер.
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Кроссинговер. Этот процесс происходит в профазе I мейоза в то время, когда гомологичные хромосомы тесно сближены в результате конъюгации и образуют биваленты. В ходе кроссинговера осуществляется обмен соответствующими участками между взаимно переплетающимися хроматидами гомологичных хромосом (рис. 3.72). Этот процесс обеспечивает перекомбинацию отцовских и материнских аллелей генов в каждой группе сцепления. В разных предшественниках гамет Кроссинговер происходит в различных участках хромосом, в результате чего образуется большое разнообразие сочетаний родительских аллелей в хромосомах.
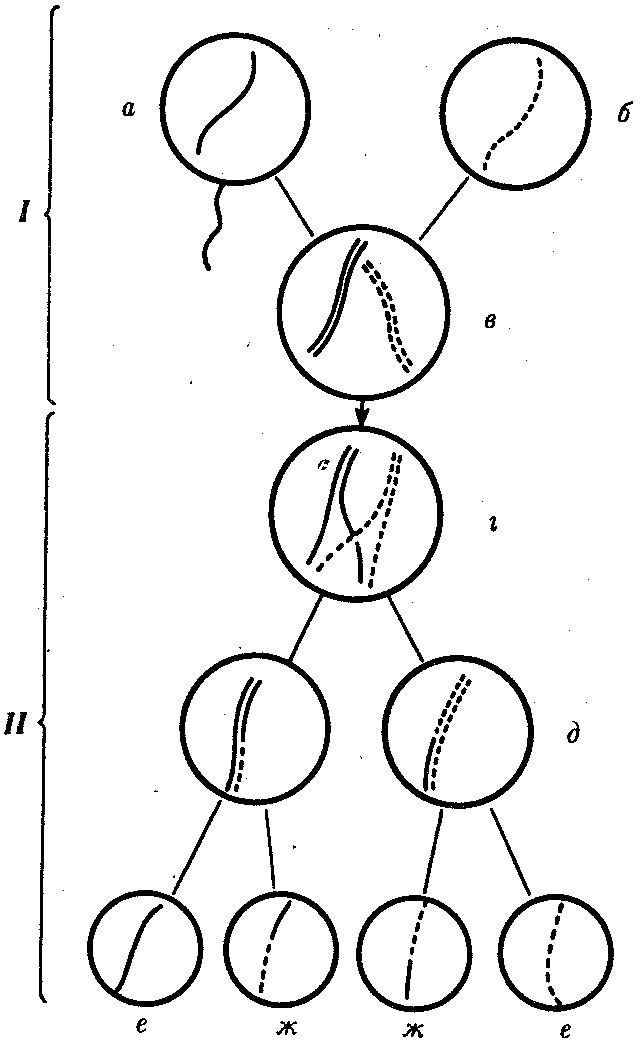
Рис. 3.72. Кроссинговер как источник генетического разнообразия гамет:
I — оплодотворение родительских гамет а и б с образованием зиготы в; II — гаметогенез в организме, развившемся из зиготы в; г — кроссинговер, происходящий между гомологами в профазе I; д — клетки, образовавшиеся после 1-го мейотического деления; е, ж — клетки, образовавшиеся после 2-го деления мейоза (е — некроссоверные гаметы с исходными родительскими хромосомами; ж — кроссоверные гаметы с перекомбинацией наследственного материала в гомологичных хромосомах)
Понятно, что кроссинговер как механизм рекомбинации эффективен лишь в том случае, когда соответствующие гены отцовской и материнской хромосом представлены разными аллелями. Абсолютно идентичные группы сцепления при кроссинговере не дают новых сочетаний аллелей.
Кроссинговер происходит не только в предшественницах половых клеток при мейозе. Он наблюдается также в соматических клетках при митозе. Соматический кроссинговер описан у дрозофилы, у некоторых видов плесеней. Он осуществляется в ходе митоза между гомологичными хромосомами, однако его частота в 10 000 раз меньше частоты мейотического кроссинговера, от механизма которого он ничем не отличается. В результате митотического кроссинговера появляются клоны соматических клеток, различающихся по содержанию в них аллелей отдельных генов. Если в генотипе зиготы данный ген представлен двумя разными аллелями, то в результате соматического кроссинговера могут появиться клетки с одинаковыми либо отцовскими, либо материнскими аллелями данного гена (рис. 3.73).

Рис. 3.73. Кроссинговер в соматических клетках:
1 — соматическая клетка, в гомологичных хромосомах которой ген А представлен двумя разными аллелями (А и а); 2 — кроссинговер; 3 — результат обмена соответствующими участками между гомологичяыми хромосомами; 4 — расположение гомологов в плоскости экватора веретена деления в метафазе митоза (два варианта); 5 — образование дочерних клеток; 6 — образование гетерозитотиых по гену А клеток, сходных с материнской клеткой по набору аллелей (Аа); 7 — образование гомозиготных по гену А клеток, отличающихся от материнской клетки по набору аллелей (АА или аа)
40.Наследование. Типы наследования. Особенности аутосомного, Х-сцепленного и голандрического типов наследования. Полигенное наследование.
под наследственностью понимают свойство клеток или организмов в процессе самовоспроизведения передавать новому поколению способность к определенному типу обмена веществ и индивидуального развития, в ходе которого у них формируются общие признаки и свойства данного типа клеток и вида организмов, а также некоторые индивидуальные особенности родителей. На популяционно-видовом уровне организации жизни наследственность проявляется в поддержании постоянного соотношения различных генетических форм в ряду поколений организмов данной популяции (вида).
Наследственность – свойство живых организмов, обеспечивающее материальную преемственность онтогенеза в определенных условиях внешней среды. Гены детерминируют последовательность полипептидной цепи.
Наследование – передача информации от одного поколения к другому. Благодаря наследственности стало возможно существование популяций, видов и других групп.
Аутосомное наследование. Характерные черты аутосомного наследования признаков обусловлены тем, что соответствующие гены, расположенные в аутосомах, представлены у всех особей вида в двойном наборе. Это означает, что любой организм получает такие гены от обоих родителей. В соответствии с законом чистоты гамет в ходе гаметогенеза все половые клетки получают по одному гену из каждой аллельной пары (рис. 6.6). Обоснованием этого закона является расхождение гомологичных хромосом, в которых располагаются аллельные гены, к разным полюсам клетки в анафазе I мейоза
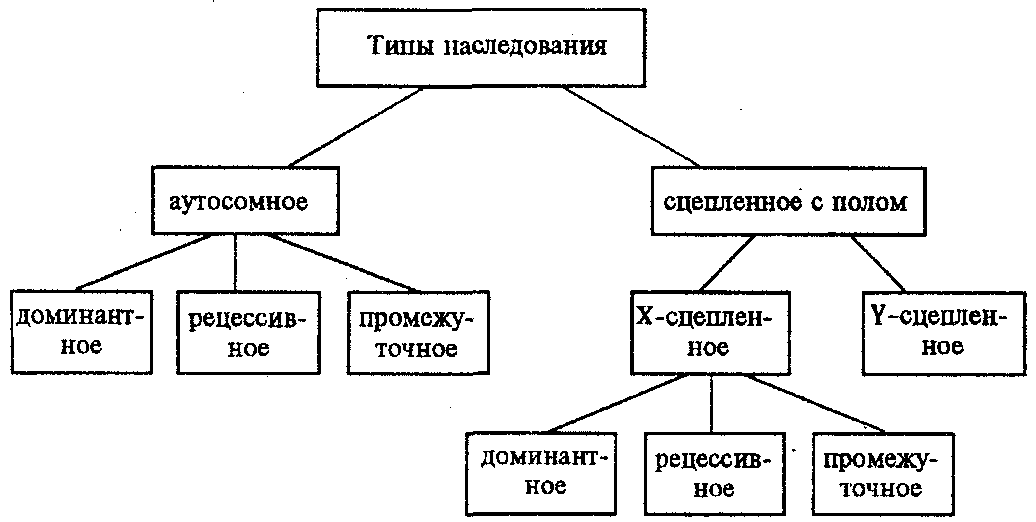
Ввиду того что развитие признака у особи зависит в первую очередь от взаимодействия аллельных генов, разные его варианты, определяемые разными аллелями соответствующего гена, могут наследоваться по аутосомно-доминантному или аутосомно-рецессивному типу, если имеет место доминирование. Возможен также промежуточный тип наследования признаков при других видах взаимодействия аллелей (см. разд. 3.6.5.2).
При доминировании признака, описанном Г. Менделем в его опытах на горохе, потомки от скрещивания двух гомозиготных родителей, различающихся по доминантному и рецессивному вариантам данного признака, одинаковы и похожи на одного из них (закон единообразия F1). Описанное Менделем расщепление по фенотипу в F2 в отношении 3:1 в действительности имеет место лишь при полном доминировании одного аллеля над другим, когда гетерозиготы фенотипически сходны с доминантными гомозиготами (закон расщепления в F2).
Наследование рецессивного варианта признака характеризуется тем, что он не проявляется у гибридов F1, а в F2 проявляется у четверти потомков.
В случаях формирования у гетерозигот нового варианта признака по сравнению с гомозиготами, что наблюдается при таких видах взаимодействия аллельных генов, как неполное доминирование, кодоминирование, межаллельная комплементация, гибриды F1 не похожи на родителей, а в F2 образуется три фенотипических группы потомков (рис. 6.7, II).
Сцепленное с полом наследование.
Х-сцепленное наследование. Х-хромосома присутствует в кариотипе каждой особи, поэтому признаки, определяемые генами этой хромосомы, формируются у представителей как женского, так и мужского пола. Особи гомогаметного пола получают эти гены от обоих родителей и через свои гаметы передают их всем потомкам. Представители гетерогаметного пола получают единственную Х-хромосому от гомогаметного родителя и передают ее своему гомогаметному потомству.
У млекопитающих (в том числе и человека) мужской пол получает Х-сцепленные гены от матери и передает их дочерям. При этом мужской пол никогда не наследует отцовского Х-сцепленного признака и не передает его своим сыновьям.
Так как у гомогаметного пола признак развивается в результате взаимодействия аллельных генов, различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. Х-сцепленный доминантный признак (красный цвет глаз у дрозофилы) передается самкой всему потомству. Самец передает свой Х-сцепленный доминантный признак лишь самкам следующего поколения. Самки могут наследовать такой признак от обоих родителей, а самцы — только от матери.
Х-сцепленный рецессивный признак, (белый цвет глаз у дрозофилы) у самок проявляется только при получении ими соответствующего аллеля от обоих родителей (XaXa). У самцов XaY он развивается при получении рецессивного аллеля от матери. Рецессивные самки передают рецессивный аллель потомкам любого пола, а рецессивные самцы —только «дочерям» (см. рис. 6.8).
Голандрическое наследование. Активно функционирующие гены Y-хромосомы, не имеющие аллелей в Х-хромосоме, присутствуют в генотипе только гетерогаметного пола, причем в гемизиготном состоянии. Поэтому они проявляются фенотипически и передаются из поколения в поколение лишь у представителей гетерогаметного пола. Так, у человека признак гипертрихоза ушной раковины («волосатые уши») наблюдается исключительно у мужчин и наследуется от отца к сыну.
Большинство количественных признаков организмов определяется полигенами, т.е. системой неаллельных генов, одинаково влияющих на формирование данного признака. Взаимодействие таких генов в процессе формирования признака называют полимерным. Оно сводится чаще всего к суммированию действия сходных аллелей этих генов, определяющих формирование одинакового варианта признака. Совместное действие полигенов обусловливает различную экспрессивность — степень выраженности признака, зависящую от дозы соответствующих аллелей. В основе появления в геноме таких генов, очевидно, лежит их дупликация или амплификация (см. разд. 3.6.4.3). Это позволяет увеличить синтез соответствующего продукта в клетках организма.
По полимерному типу взаимодействия у человека определяется интенсивность окраски кожных покровов, зависящая от уровня отложения в клетках пигмента меланина. В геноме человека имеется четыре гена, отвечающих за этот признак. В генотипе все они представлены в двойной дозе. В связи с тем что эти гены отвечают за развитие одного и того же признака, их обозначают одной и той же буквой, но с разными символами, чтобы подчеркнуть их неаллельность — P1P2P3P4.
Наличие в генотипе восьми доминантных аллелей в системе полигенов, определяющих цвет кожи, обусловливает максимальную ее пигментацию, наблюдаемую у африканских негров (P1P1P2P2P3P3P4P4). Полное отсутствие доминантных аллелей у. рецессивных гомозигот (р1р2р2р3р3р4р4) проявляется в виде минимальной пигментации у европеоидов. Большее или меньшее количество доминантных аллелей, колеблющееся от 8 до 0, обеспечивает разную интенсивность окраски кожи (рис. 3.80). Полимерное взаимодействие генов лежит в основе определения главным образом количественных признаков (рост, масса организма, возможно, интеллект).
41.Количественная и качественная специфика проявления генов в признаках: пенетрантность, экспрессивность, плейотропность, генокопии.
Никакие признаки не наследуются. Признаки развиваются на основе взаимодействия генотипа и среды. Наследуется только генотип, т.е. комплекс генов, который определяет норму биологической реакции организма, изменяющую проявление и выраженность признаков в разных условиях среды. Таким образом, организм реагирует на свойства внешней среды. Иногда один и тот же ген в зависимости от генотипа и от условий внешней среды по-разному проявляет признак или меняет полноту выраженности.
Степень проявления фенотипа – экспрессивность. Образно ее можно сравнить со степенью тяжести болезни в клинической практике. Экспрессивность подчиняется законам распределения Гаусса (некоторые в малом или среднем количестве). В основе изменчивости экспрессивности лежат и генетические факторы, и факторы внешней среды. Экспрессивность – очень важный показатель фенотипического проявления гена. Количественно ее степень определяют, используя статистический показатель.
Экспрессивность также является показателем, характеризующим фенотипическое проявление наследственной информации. Она характеризует степень выраженности признака и, с одной стороны, зависит от дозы соответствующего аллеля гена при моногенном наследовании или от суммарной дозы доминантных аллелей генов при полигенном наследовании, а с другой — от факторов среды. Примером служит интенсивность красной окраски цветков ночной красавицы, убывающая в ряду генотипов АА, Аа, аа, или интенсивность пигментации кожи у человека, увеличивающаяся при возрастании числа доминантных аллелей в системе полигенов от 0 до 8 (см. рис. 3.80). Влияние средовых факторов на экспрессивность признака демонстрируется усилением степени пигментации кожи у человека при ультрафиолетовом облучении, когда появляется загар, или увеличением густоты шерсти у некоторых животных в зависимости от изменения температурного режима в разные сезоны года.
Генетический признак может даже не проявляться в некоторых случаях. Если ген есть в генотипе, но он вовсе не проявляется – он пенетрирован. (русский ученый Тимофеев-Рисовский 1927 год). Пенетрантность – количество особей (%), проявляющих в фенотипе данный ген, по отношению к количеству особей, у которых этот признак мог бы проявиться. Пенетрантность свойственна проявлению многих генов. Важен принцип – «все или ничего» - либо проявляется, любо нет.
- наследственный панкреатит – 80%
- вывих бедра – 25%
- пороки развития глаз
- ретинобластома – 80%
- отосклероз – 40%
колотокома – 10%
Пенетрантностъ отражает частоту фенотипического проявления имеющейся в генотипе информации. Она соответствует проценту особей, у которых доминантный аллель гена проявился в признак, по отношению ко всем носителям этого аллеля. Неполная пенетрантность доминантного аллеля гена может быть обусловлена системой генотипа, в которой функционирует данный аллель и которая является своеобразной средой для него. Взаимодействие неаллельных генов в процессе формирования признака может привести при определенном сочетании их аллелей к непроявлению доминантного аллеля одного из них.
Хорея Гентингтона проявляется в непроизвольном подергивании головы. Конечностей, постепенно прогрессирует и приводит к смерти. Может проявиться в раннем постэмбриональном периоде, в зрелом возрасте или не проявиться вообще. И экспрессивность, и пенетрантность поддерживаются естественным отбором, т.е. гены, контролирующие патологические признаки могут иметь разную экспрессивность и пенетрантность: заболевают не все носители гена, а у заболевших степень проявления будет различна. Проявление или неполное проявление признака, а так же его отсутствие зависит от среды и от модифицирующего действия других генов.
Ген может действовать плейотропно (множественно), т.е. опосредовано влиять на течение разных реакций и развитие многих признаков. Гены могут оказывать влияние на другие признаки на разных стадиях онтогенеза. Если ген включается в позднем онтогенезе, то оказывается незначительное действие. Если на ранних стадиях – изменения более значительны.
Фенилкетанурия. У больных есть мутация, которая выключает фермент – фенилаланин – гидролазу. Поэтому фенилаланин не превращается в тирозин. В результате в крови количество фенилаланина повышается. Если выявить эту патологию рано (до 1 месяца) и перевести ребенка на другое питание, развитие идет нормально, если позднее – понижен размер головного мозга, умственная отсталость, не развиваются нормально, отсутствует пигментация, умственные способности минимальны.
Плейотропность отражает интеграцию генов и признаков.
У человека есть патологический ген, приводящий к синдрому Фанкони (порок развития или отсутствие большого пальца, порок или отсутствие лучевой кости, недоразвитие почки, коричневые пигментные пятна, нехватка кровяных телец).
Есть ген, связанный с Х-хромосомой. Невосприимчивость к инфекциям и нехватка кровяных телец.
Доминантный ген, сцепленный с Х-хромосомой – пилонефрит, лабиринтная тугоухость.
Синдром Марфани – паучьи пальцы, вывих хрусталика глаза, пороки развития сердца.
Генокопия (греч. genos род, происхождение + лат. copia множество) - термин в 1957 г. предложил немецкий генетик Нахтсхейм (H.Nachtsheim).Обозначает сходные изменения одного и того же признака под влиянием разных неаллельных генов, которые иногда называют миметическими генами гетерогенной группы.
genocopies - генокопии.
Oдинаковые изменения фенотипа, обусловленные аллелями разных генов, а также имеющие место в результате различных генных взаимодействий или нарушений различных этапов одного биохимического процесса с прекращением синтеза конечного продукта, - например, у Drosophila melanogaster известен ряд мутаций неаллельных генов, обусловливающих фенотип “красные глаза” (нарушен синтез коричневого пигмента).
42.Изменчивость. Формы изменчивости: модификационная и генотипическая, их значение в онтогенезе и эволюции.
Изменчивость
Изменчивость – свойство живых организмов существовать в разных формах. Групповая и индивидуальная изменчивость – классификация по эволюционному значению. Изменчивость, реализованная группой организмов, называется групповой, у одного организма или группы его клеток – индивидуальная.
По характеру изменения признаков и механизму:
--фенотипическая
- случайная
- модификационная
--генотипическая
- соматическая
- генеративная (мутационная, комбинативная)
а) генная
б) хромосомная
в) геномная
Модификационная изменчивость отражает изменение фенотипа под воздействием факторов внешней среды (усиление и развитие мышечной и костной массы у спортсменов, увеличение эритропоэза в условиях высокогорья и крайнего севера).
43.Фенотипическая изменчивость и её виды. Модификации и их характеристики. Норма реакции признака. Фенокопии. Адаптивный характер модификаций.
По характеру изменения признаков и механизму:
--фенотипическая
- случайная
- модификационная
Модификационная изменчивость отражает изменение фенотипа под воздействием факторов внешней среды (усиление и развитие мышечной и костной массы у спортсменов, увеличение эритропоэза в условиях высокогорья и крайнего севера). Частный случай фенотипической изменчивости – фенокопии. Фенокопии – вызванные условиями внешней среды фенотипические модификации, имитирующие генетические признаки. Под влиянием внешних условий на генетически нормальный организм копируются признаки совсем другого генотипа. Проявление дальтонизма может произойти под влиянием питания, плохой психической конституции, повышенной раздражительности. У человека возникает заболевание витилиго (1% людей) – нарушение пигментации кожи. Генетический дефект есть у 30% болеющих, у остальных – профессиональное витилиго (воздействие на организм особых химических и отравляющих веществ). В Германии 15 лет назад рождались дети с фекомелией – укороченными ластовидными руками. Выяснилось. Что рождение таких детей происходило, если мать принимала Телидомид (успокоительное средство, показанное беременным). В результате нормальный немутантный генотип получал мутацию.
Фенокопии появляются в большинстве случаев при действии внешней среды на ранних стадиях эмбриогенеза, что приводит к врожденным заболеваниями порокам развития. Наличие фенокопий затрудняет диагностику заболеваний.