

130
140 150
20 30
40 50 60 70 80___90
100 ПО
40
Тигр: х-.'-':'":':!
современный ареал
Орангутан
|!
Бабирусса Слон
Утка-мандаринка
.'.'•и
Тапирчепрачный 1000 0 1000км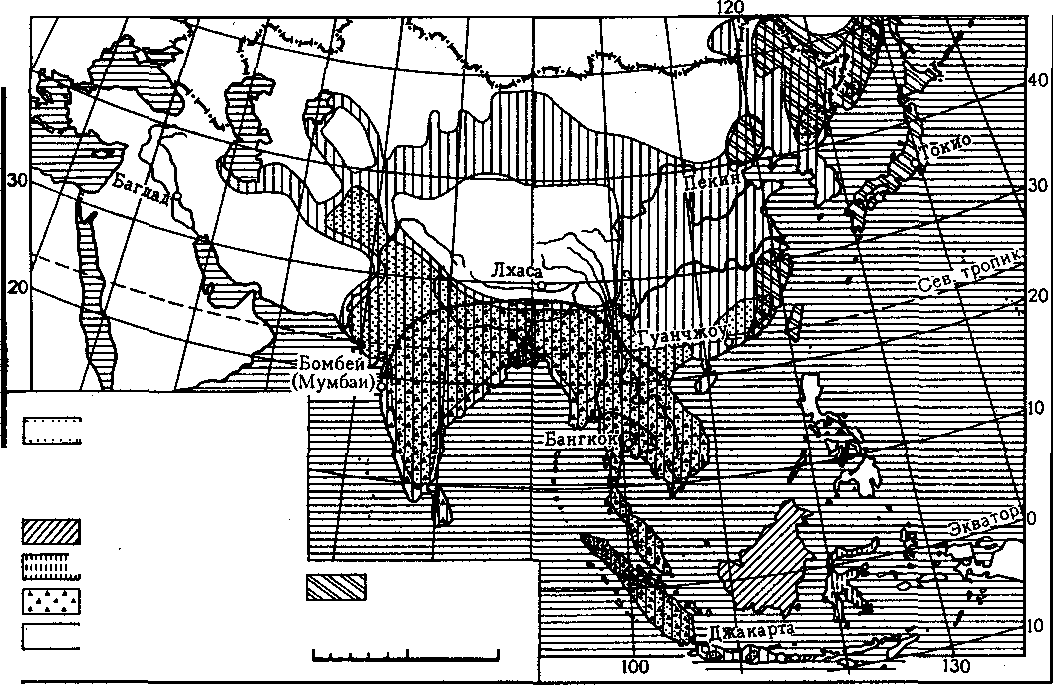
1[|[|[| Примерная граница ц-111" распространения в середине XIX в.
Рис. 22. Распространение некоторых животных в зарубежной Азии
Острова Сулавеси и Малые Зондские в зоологическом отношении занима-|ют особое место. К числу эндемичных животных Сулавеси относятся дикая ^свинья бабирусса (ВаЪугозза ЬаЬугима), карликовый буйвол аноа (ВиЬаШз >рге551сотю) и черный макак, а к представителям австралийской фауны — ^сумчатые кускусы, большеногие куры и многие другие птицы.
В особую Индийскую подобласть выделяют Индию, Шри-Ланку и Индокитай. В составе фауны этой подобласти наряду со многими типичными представителями Индо-Малайской области присутствуют выходцы из Эфиопской области и Голарктики. Животный мир Индийской подобласти отличается видовым многообразием и большой численностью особей. Особенно это относится к Индии, где убийство любых живых существ запрещено религией, поэтому здесь очень редко истребляют даже вредных животных.
В фауне Индии и Индокитая характерно присутствие индийского слона. Дикие слоны до сих пор еще встречаются в малонаселенных районах предгорий Гималаев, в лесах Шри-Ланки и других местах. Домашний слон, приученный исполнять трудные и сложные работы,— одно из самых типичных животных Индии и стран Индокитая.
Местное население приручает также дикого быка — гаура (гаяла). Одомашнен и широко распространен как рабочий скот индийский буйвол, В густых приречных зарослях часто встречается дикий индийский кабан. В тех районах, где сохранились значительные массивы лесов, обитают крупная антилопа ниль-гау (Возе1арНш 1га§осате1из) и четырехрогая антилопа (Те1гасегиз диа^псогтз), олени мунтжак и аксис (Сеп>из ах&) — один из самых красивых представителей этого семейства, живущий в лесных районах, богатых водой. Из хищников
117
распространены тигр, леопард и особая форма леопарда — черная пантера, наносящие значительный ущерб скотоводству. В пределах пустыни Тар изредка встречается лев, проникший сюда из Эфиопской области.
Для Индии и Индокитая характерно обилие обезьян, которые распространены повсюду: в лесах, саваннах, садах, вблизи населенных пунктов и даже в городах. Они поедают плоды и портят посевы, нанося большой ущерб населению. В Индии встречаются собакообразные обезьяны, в Индокитае — гиббоны, макаки и др. В пределах подобласти как в лесах, так и вблизи человеческого жилья обитают полуобезьяны, или лемуры. Для Индокитая, так же как и для островов, характерны шерстокрылы.
Настоящее бедствие для местного населения — обилие различных пресмыкающихся, особенно ядовитых змей, от укусов которых ежегодно гибнут тысячи людей. В водах Ганга и других крупных рек водятся гигантские крокодилы (ОашаНз §ап§е(1си5), достигающие 6 м длины.
Яркостью оперения и разнообразием форм поражает мир птиц. Среди них — обыкновенный павлин (Раио сп$Шш), фазан, виды диких кур, от которых происходят домашние породы, различные дрозды и т.д. Из насекомых особенно много различных пестро окрашенных бабочек, гигантских пауков-птицеедов, питающихся мелкими птицами. В Индии водится дикая пчела — родоначальница домашней пчелы.
Прямое уничтожение ценных видов растений и животных (охота, браконьерство, незаконная торговля), а главное — изменение их местообитаний как результат антропогенного воздействия, привели к тому, что многие виды фауны Евразии оказались под угрозой исчезновения. Это 471 вид млекопитающих, 389 видов птиц, 276 видов рыб, 85 видов пресмыкающихся и 33 вида земноводных. Разрушено около двух третей всех мест обитания диких животных Азии. В Китае, одной из 12 стран мира, отличающихся «мегабиоразнообрази-ем», под угрозой исчезновения находятся 15—20% видов. Из семи эндемич-ных видов млекопитающих Западной Азии четыре (аравийский леопард, полосатая гиена, аравийский тар и аравийский волк) находятся под угрозой исчезновения. Практически не улучшается ситуация с потерей видов и их местообитаний в Западной Европе.
ЧЕЛОВЕК
В составе фауны Евразии есть представители обоих подотрядов отряда приматов: полуобезьяны и человекоподобные приматы, в том числе современные человекообразные обезьяны. В пределах Евразии были обнаружены ископаемые останки древнейших гоминид — предков человека современного вида. В разных частях материка найдены костные останки предков человека различных стадий развития. Эти факты свидетельствуют о том, что Евразия входила в область становления и развития человека.
В южной части Евразии обнаружены ископаемые останки человекообразных обезьян, живших в конце неогена. В начале четвертичного периода (в виллафранке) в Передней, Южной и Юго-Восточной Азии, судя по некоторым находкам, возможно, жили высшие приматы, родственные австрало-
118
[питекам Африки. Однако достоверность этих данных до сих пор не подтверж-Гдена.
Бесспорны и довольно многочисленны в Евразии находки архантропов,
соторых относят к первой стадии в развитии собственно человека. Останки ар-[хантропов, живших 800—500 тыс. лет назад, были обнаружены на Яве. Там же [найдены костные останки архантропов более позднего времени, соответству-[ющего миндельскому (окскому) ледниковые, т.е. нижнему плейстоцену (абсо-[лютный возраст — 500 тыс. лет). Это первая находка древнейшего человека,
получившего впоследствии название прямоходящего питекантропа (РШгесап1горш
Архантропы были широко распространены не только на юго-востоке Евра-|зии. Находки, почти одновозрастные с питекантропом, обнаружены также во многих районах субтропического и умеренного поясов с относительно мягким 1в то время климатом — на западе Европы (гейдельбергский человек, относи-[мый к понцу или гюнц-минделю, т.е. тоже к нижнему плейстоцену), в Центральной Европе (питекантроп Венгрии, сходный с яванским). Архантропы оби-[тали также на Средиземноморском побережье Европы и Азии. Районы Восточ-[ной Европы и Северной Азии, отличавшиеся суровым континентальным кли-I матом и покрывавшиеся льдами, начали заселяться гораздо позднее. Наиболее [поздний по абсолютному возрасту представитель архантропов в Евразии — [синантроп, ископаемые останки которого обнаружены недалеко от Пекина ! (рис. 23).
Широкое распространение получили в Евразии палеоантропы. Их существование было очень продолжительным — от среднего до позднего плейстоцена. .Останки палеоантропов найдены в Западной Европе, включая Британские ос-юва, в Центральной и Передней Азии, на Яве. В Передней Азии в пещерах горы Кармель на территории современного Израиля наряду с остатками типичных палеоантропов были найдены скелеты переходного типа между палео-антропами и неоантропами. Поздние палеоантропы, или неандертальцы, возможно, образовали непрогрессивную ветвь, завершившую свое существование во второй половине позднего плейстоцена. Эволюция группы ранних палеоантропов, обитавших в восточной части Средиземноморья и обладавших прогрессивными чертами, привела в дальнейшем к возникновению современного человека.
Неоантропы, по своим антропологическим особенностям практически не отличавшиеся от современного человека, появились в Европе и Юго-Восточной Азии примерно 40—38 тыс. лет назад, во второй половине верхнего плейстоцена, что соответствует верхнему палеолиту. Европейские люди современного типа получили название кроманьонцев от грота Кро-Маньон в Центральном массиве Франции, где были обнаружены их скелетные останки. Расселяясь по материку, они проникали на север и восток, в пределы Восточной Европы и Сибири. По мере приспособления к разнообразной и резко менявшейся природной среде, а также в результате контактов и смешения формировались современные расы и антропологические типы Евразии.
Для Евразии характерны исключительная сложность и разнообразие расового состава населения. При этом многие расовые подразделения являются автохтонными. Евразия (точнее, ее восточная часть) — родина монголоидов. В южной части материка сформировалась океанийская ветвь экваториальной расы.
к западу от Гринвича
0 к востоку от Гринвича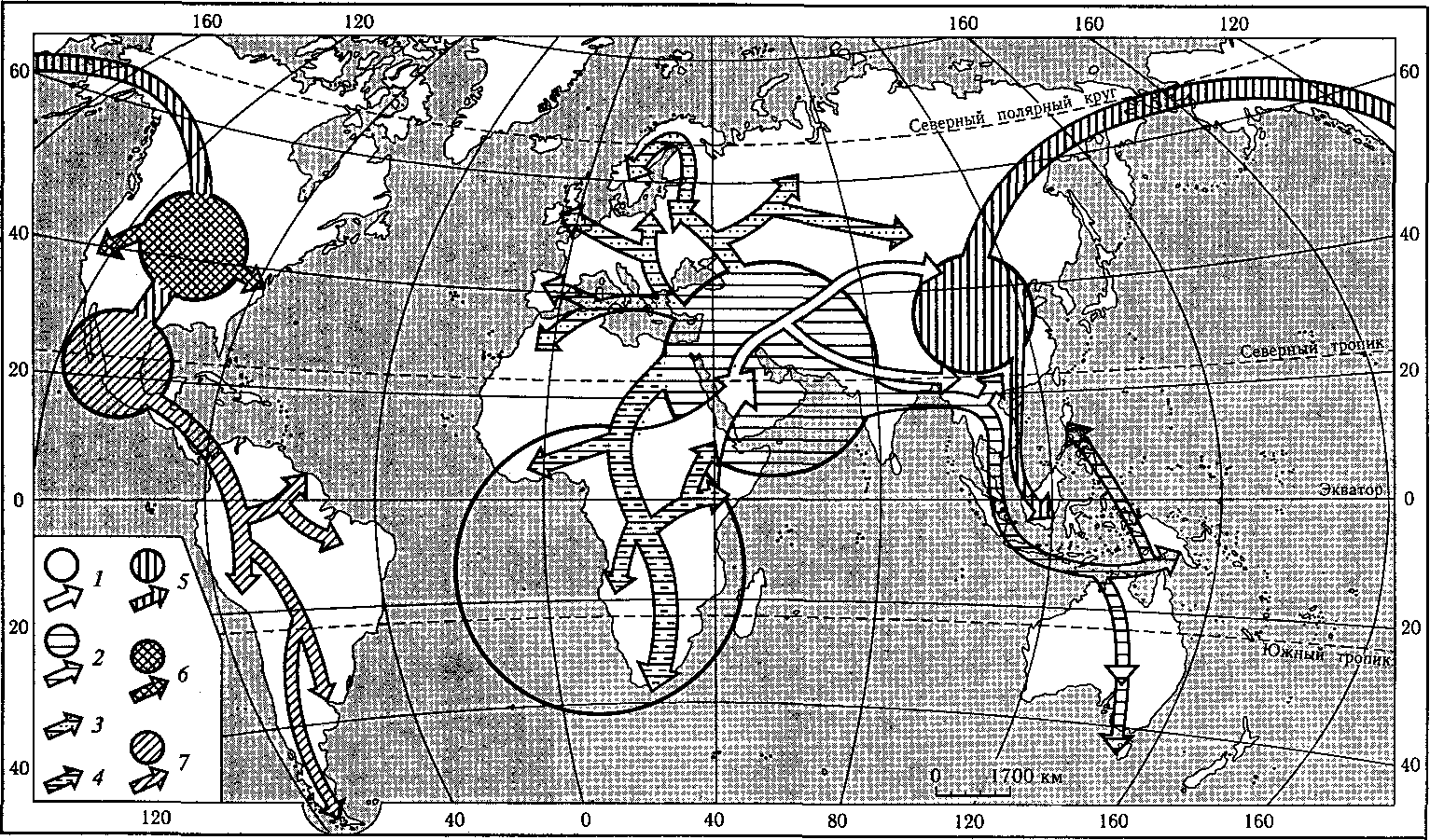
Рис. 23. Центры становления человека и пути его расселения (по В. П.Алексееву):
1 — прародина человечества и расселение из нее; 2 — первичный западный очаг расообразования и расселение протоавстралоидов; 3 —
расселение протоевропеоидов; 4 — расселение протонегроидов; 5 — первичный восточный очаг расообразования и расселение протоамерика-
ноидов; 6 — североамериканский третичный очаг и расселение из него; 7 — центрально-южноамериканский очаг и расселение из него
Вместе с Северной Африкой юго-западная часть Евразии была родиной юж-[вых европеоидов, а северная ветвь европеоидной расы образовалась в пределах Европы. В Евразии происходила дифференциация больших рас на расы второго порядка и шел процесс образования большого количества антропологических типов. Огромное значение для расообразования в различные периоды имело смешение и образование переходных и смешанных рас.
Дифференциация рас в пределах огромного Евразийского материка — ре-. зультат, во-первых, тех изменений, которые претерпевали европеоиды в ходе [их продвижения на север и северо-восток вслед за отступавшими ледниками [последнего оледенения; во-вторых, сложных контактов между европеоидами [и монголоидами, а также между теми и другими, с одной стороны, и пред-(ставителями океанийской ветви экваториальной расы — с другой.
Юго-Западная Азия и южная половина Европы, включая Среднюю Евро-
, являются не только районами давнего заселения, но вместе с Северной Африкой входят также в ареал формирования древней ветви европеоидов — *южной европеоидной, или средиземноморской, расы. Характерные признаки ;ее — смуглый цвет кожи, темный цвет волос и глаз, длинноголовость, невысокий рост. Наиболее полно признаки южных европеоидов выражены у насе-[ления Западного Средиземноморья.
Представители южных европеоидов в Центральной Азии — горные таджики, по мнению антропологов, могут рассматриваться как характерные, «чистые» представители большой европеоидной расы. У них преобладают темные волосы и глаза, брахикефалия, резко выступающий нос. Этот антропологический тип выделяется под названием памирского.
Заселение северных и северо-западных районов Европы стало возможным только после исчезновения ледниковых покровов. Смуглые, темноволосые и темноглазые европеоиды при продвижении на север, в районы сурового климата с четко выраженной сезонностью, постепенно утрачивали значительную часть темного пигмента. Процесс депигментации — один из важнейших факторов формирования северной ветви европеоидов, он происходил очень медленно, постепенно, без четких границ и переходов как во времени, так и в пространстве. Поэтому между крайними типами южной темноокрашенной ветви европеоидов (средиземноморской) и северной (балтийской), со светлой кожей, русыми волосами и светлыми глазами, располагается полоса распространения смешанных и переходных типов, к которым принадлежит основная часть населения Европы.
Монголоидная раса, формировавшаяся в восточных районах Центральной Азии, по мере распространения на север, восток и юг испытывала значительную дифференциацию. Еще в самом начале образования большой монголоид-ной расы из нее выделилась американская ветвь, давшая начало коренному населению Америки. Для представителей азиатской ветви монголоидов, становление которых проходило в условиях сухого и жаркого климата Центральной Азии, характерны желтовато-смуглая кожа, прямые и жесткие черные волосы, широкое лицо, широкий нос с низким переносьем и открытыми ноздрями, особое строение верхнего века с защитной складкой — эпикантусом. Рост азиатских монголоидов небольшой, а скелет довольно массивный. Люди с наиболее полно выраженными монголоидными признаками образуют северо-монголоидную, или центрально-азиатскую, расу.
121
Большая часть внутренних районов Евразии населена народами, антропологические особенности которых сложились в результате контактов между европеоидами и монголоидами как на ранних этапах расообразования, так и в средневековье и в новое время. К числу древних контактных форм принадлежит уральская раса. Она распространена в равнинных и горных районах Западной Сибири и не проникает на восток дальше Енисея. В большинстве регионов Евразии, за исключением крайнего востока, первоначально преобладало европеоидное население, монголоидные черты были привнесены в процессе распространения на запад северной ветви монголоидов. Поэтому у современного населения монголоидные черты получают преимущественное значение при движении на восток, европеоидные — при движении на запад.
Особое место среди населения Северной Европы занимают лопари, населяющие самые северные районы некоторых стран. У них отчетливо выражены некоторые монголоидные признаки: низкий рост, темная пигментация, круглая голова, иногда даже типичное для монголоидов строение века. Очевидно, эти черты связаны со сравнительно поздним монголоидным влиянием, результаты которого не успели исчезнуть в условиях географической среды полярных районов.
В Южной Азии (на юге Индокитая или Индостана) сформировалась океанийская ветвь экваториальной расы, распространившаяся затем на юго-восток и юг — в Австралию и Океанию и частично на северо-восток. В настоящее время черты этой ветви четко выражены у некоторых темнокожих и малорослых народов, живущих в Центральной Индии и на Шри-Ланке. Древние контакты между океанийской ветвью и южными европеоидами в Южной Азии привели к образованию южноиндийской переходной расы, к которой принадлежат, например, сингалы Шри-Ланки или некоторые народы южной части Индостана.
На Андаманских островах, во внутренних районах полуострова Малакка, на острове Лусон в Филиппинском архипелаге обитают немногочисленные низкорослые племена, часто объединяемые в одну группу негритосов, или азиатских пигмеев. Хотя они и имеют некоторые черты внешнего сходства с африканскими негриллями, обусловленные общностью условий местообита-ния, генетически эти разобщенные группы малорослых народов Азии и Африки не связаны между собой. Очевидно, азиатские пигмеи, как и африканские, сформировались из более древних высокорослых форм под влиянием изменившейся природной среды, что могло быть обусловлено переселением их предков во влажные тропические леса из других районов Юго-Восточной Азии.
Большое значение для формирования антропологических особенностей населения Юго-Восточной Азии имеют взаимоотношения между океанийской ветвью экваториальной расы и монголоидами. Последние распространялись из первоначального центра своего возникновения не только на север, но и на юг. При этом значительно изменялись антропологические особенности: исчезал эпикантус как признак, не нужный в условиях Юго-Восточной Азии, происходила общая грацилизация скелета (утонынение костей, уменьшение длины тела), несколько усиливалась пигментация. В результате этих изменений и смешения с океанийской ветвью происходило образование расового типа, существенным образом отличавшегося от северных и восточных монголоидов. По-
122
:пенно сложилась особая южно-монголоидная, или азиатско-тихоокеанская, 1са, которую большинство антропологов считают переходной между монго-юидной и экваториальной большими расами. К ней принадлежит население >жных районов Китая, значительная часть народов Индокитайского полуост->ва и островов Малайского архипелага. Иногда к этой расе относят также юнцев, в других случаях их выделяют как особый расовый тип на границе гежду монголоидами и океанийской ветвью.
к
востоку от
Гринвича
0 к западу от Гринвича
40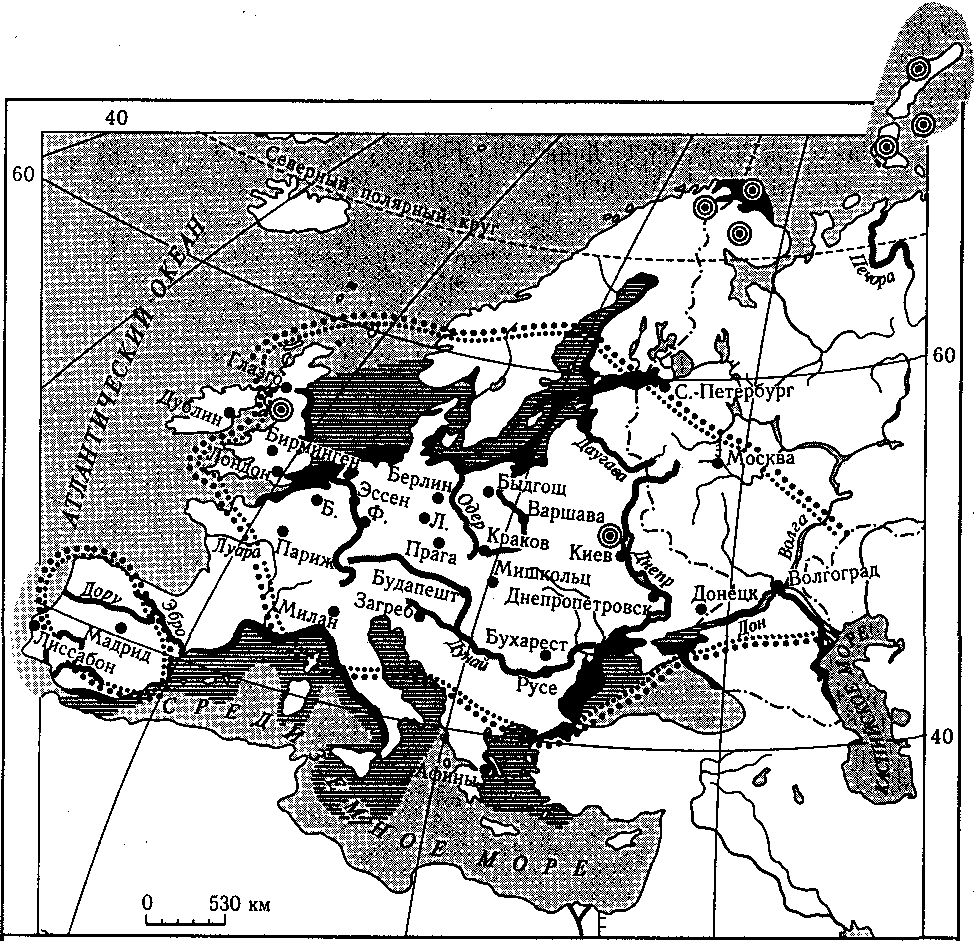
..V; Кислотные дожди
•и» Загрязненные реки
|) Радиоактивное загрязнение
Буквами на карте обозначены:
Загрязнение моря Сильное загрязнение моря
Города с сильно загрязненным воздухом
чены: Б,— Брюссель Ф. — Франкфурт-на-Майне ' Л.—Лейпциг