

Свойства бинокулярного восприятия:
Фузия – слияние полностью идентичных изображений двух сетчаток. При этом объект воспринимается двухмерным, плоскостным.
Слияние двух неидентичных изображений сетчаток. При этом объект воспринимается объемно, трехмерно.
Соперничество полей зрения. От правой и левой сетчатки поступают два разных изображения. Мозг не может совместить два разных изображения, и поэтому они воспринимаются поочередно.

Остальные точки сетчатки – диспаратные. Степень диспаратности и будет определять, воспринимается ли объект трёхмерно или он будет восприниматься при соперничестве полей зрения. Если диспаратность невелика, то изображение воспринимается трёхмерно. Если диспаратность очень высокая, то объект не воспринимается.
Такие нейроны обнаружены не в 17-м, а в 18-м и 19-м полях.
Чем отличаются рецептивные поля таких клеток: для таких нейронов в зрительной коре рецептивные поля либо простые, либо сложные. В этих нейронах наблюдается различие рецептивных полей от правой и левой сетчатки. Диспаратность рецептивных полей таких нейронов может быть либо вертикальной, либо горизонтальной (см. след. страницу):
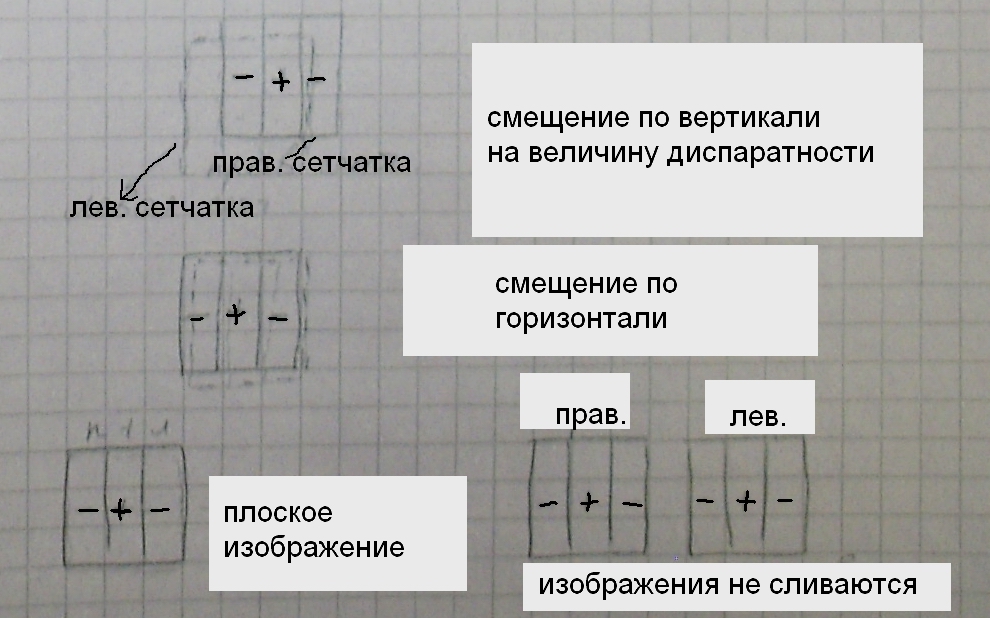
Это свойство позволяет лучше адаптироваться.
(+) Зрительная кора не позволяет говорить о том, что в ней формируется зрительный образ, то константность отсутствует во всех областях зрительной коры.
Ассоциативные области коры мозга
Где были обнаружены свойства, которыми не обладает зрительная кора (целостность и константность): заднетеменная и нижневисочная кора.
Часть волокон, идущих от сетчатки, идёт к структуре, которая называется верхнее двухолмие. Это слоистая структура (6 слоев), в которой расположены нейроны, обеспечивающие управление саккадическими движениями глаз (перевод взгляда с одного объекта на другой). Информация от клеток верхнего двухолмия идет в заднетеменную кору.
В заднетеменной коре расположены нейроны, которые имеют рецептивные поля и связаны не с изображениями на сетчатке, а с участками внешнего зрительного пространства, т.е. рецептивные поля, в которых отражается внешнее зрительное пространство.
К этим нейронам поступает информация от верхнего двухолмия (о движениях глаз) и от зрительной коры (от …).
Эта область обеспечивает все ориентационные представления субъекта, т.е. расположение объектов вокруг субъекта и расположение самого субъекта относительно этих объектов.
Эта часть заднетеменной коры называется система «Где». С помощью нейронов этой области определяется расположение субъекта и определение его предметного окружения. Эта система имеет мощные анатомические связи со зрительной корой и с верхним двухолмием. Она посылает свои эфференты в префронтальную кору. Слияние внешней информации с переработанной информацией, которая даёт зрительной коре представление об относит.форм.предметов, обеспечивает константность изображения и зрительного поля.
Нижневисочная кора имеет такие же мощные анатомические связи со зрительной корой. В этой области находятся нейроны, которые обеспечивают категоризацию объектов, т.е. разделение на определённые классы, и соответственно, опознание объектов. Эта система называется система «Что», т.е. эти нейронные обеспечивают целостность восприятия.
Эфференты системы «Что» также направляются в префронтальную кору.
Поскольку гиппокамп связан с фиксацией следов памяти, в том числе и зрительных образов, то и категоризация связана с эталонами памяти. Функция этих областей связана со зрением.
И константность, и целостность зрительного восприятия формируется в ассоциативных областях: заднетеменной и нижневисочной.
ФАЙЛ 8
Свойства корковый нейронов
Это свойство пластичности присуще первичной зрительной коре, но это свойство ограничено чувствительным или сенситивным периодом у млекопитающих.
Детекторы формируются прижизненно в
зависимости от воздействия окружающей
среды. У котят – около 6 недель, у детей
– 18 месяцев. В этот период формируются
нейроны первичной зрительной коры (![]() …).
Распознавание в зрительной коре
характеристике образа называют подобразом
(гештальтом), на уровне зрительной коры
не обнаружено одного из важных свойств
зрения – константности. В 1950-х гг.
большинство учёных считали, что
большинство органов чувств связано с
ограниченным участками сенсорной коры,
а вся остальная кора обеспечивает так
называемые ассоциации. Накапливающие
позднее экспериментальные данные
заставили изменить эту точку зрения,
поскольку для зрительной сенсорной
системы кора, связанная с анализом
информации, не ограничивается первичной
зрительной корой. У млекопитающих с
развитым зрением и приматов с переработкой
зрительной информации связаны большие
участки затылочной и височной долей. У
макаков идентифицировано от 24 до 30
отдельных экстрастриарных участков
зрительной коры. Многие из этих участков
содержат карты поля зрения, связанные
с различными характеристиками зрительных
сигналов. Основной зрительный путь
поступает в зрительную кору от сетчатки
через коленчатое тело. Другой путь идёт
в верхние бугры четверохолмия. В верхних
буграх четверохолмия зрительная карта
связана с контролем движения глаз и
направлением взгляда. Эта структура
состоит из 7 слоев клеток, они связаны
не только со зрением. Сенсорный к верхнему
двухолмию образует три карты:
зрительную, соматосенсорную и слуховую,
расположенные одна над другой. Зрительная
карта формируется частично входом от
сетчатки и частично из зрительной коры.
Эта карта организована по другом
принципу, чем в зрительной коре. Она
представляет визуальное пространство
вокруг животного. Другие сенсорные
карты: соматосенсорная и слуховая –
формируются по такому же принципу, что
и зрительная, и располагаются точка в
точку одна под другой.
…).
Распознавание в зрительной коре
характеристике образа называют подобразом
(гештальтом), на уровне зрительной коры
не обнаружено одного из важных свойств
зрения – константности. В 1950-х гг.
большинство учёных считали, что
большинство органов чувств связано с
ограниченным участками сенсорной коры,
а вся остальная кора обеспечивает так
называемые ассоциации. Накапливающие
позднее экспериментальные данные
заставили изменить эту точку зрения,
поскольку для зрительной сенсорной
системы кора, связанная с анализом
информации, не ограничивается первичной
зрительной корой. У млекопитающих с
развитым зрением и приматов с переработкой
зрительной информации связаны большие
участки затылочной и височной долей. У
макаков идентифицировано от 24 до 30
отдельных экстрастриарных участков
зрительной коры. Многие из этих участков
содержат карты поля зрения, связанные
с различными характеристиками зрительных
сигналов. Основной зрительный путь
поступает в зрительную кору от сетчатки
через коленчатое тело. Другой путь идёт
в верхние бугры четверохолмия. В верхних
буграх четверохолмия зрительная карта
связана с контролем движения глаз и
направлением взгляда. Эта структура
состоит из 7 слоев клеток, они связаны
не только со зрением. Сенсорный к верхнему
двухолмию образует три карты:
зрительную, соматосенсорную и слуховую,
расположенные одна над другой. Зрительная
карта формируется частично входом от
сетчатки и частично из зрительной коры.
Эта карта организована по другом
принципу, чем в зрительной коре. Она
представляет визуальное пространство
вокруг животного. Другие сенсорные
карты: соматосенсорная и слуховая –
формируются по такому же принципу, что
и зрительная, и располагаются точка в
точку одна под другой.
В глубоких слоях четверохолмия образуется ещё одна карта – моторная. К ней из сенсорных карт поступает информация для контроля саккадических движений глаз, направляющих взгляд на важные части зрительного поля. В реализации этой функции принимает участие и фронтальная кора, т.е. лобное глазное поле. Волокна из верхних бугров четверохолмия через ядро подушки поступают в экстрастриарную зрительную кору. Подушка – это крупное ядро, лежит перед верхним бугром четверохолмия и образует заднюю часть таламуса. Это ядро направляет волокна в экстрастриарную кору и получает от неё афференты. Экстрастриарная кора получает афференты одновременно из стриарной коры и из подушки. Благодаря сложным связям зрительных областей обеспечивается целостная деятельность зрительной системы. При передаче информации от наружного коленчатого тела к стриарной коре крупноклеточные (М-типа) волокна несут сигналы о движении и форме, а парвоцеллюлярные (P-волокна) – о цвете и частично о форме. Причём эти волокна изолированы друг от друга и при входе в кору. Магноцеллюлярные волокна идут к клеткам, выделяющим края, а парвоцеллюлярные волокна – к клеткам с двойной цветовой оппонентностью и к клеткам, выделяющим положение контуров стационарных объектов.
Таким образом, в зрительной системе имеется три потока информации:
О форме, движении и стереоскопии;
О цвете;
Обо всех четырех характеристиках зрительного окружения.
Такие три раздельные потока информации
поступаю и в экстрастриарную кору (поля
![]() ).
Это 18-е поле Бродмана. Зрительные путь
прослежены до 19-го поля, содержащего
области
).
Это 18-е поле Бродмана. Зрительные путь
прослежены до 19-го поля, содержащего
области
![]() и
и
![]() ,
и дальше во многие другие части коры, в
том числе, теменной и височной. Таких
участков в коре насчитывают до 30 и они
связаны между собой волокнами в обоих
направлениях. Информация из стриарной
коры направляется в другие части мозга,
в т.ч. в базальные ядра и в промежуточный
мозг. Особенность: они связаны со
зрительной корой и относятся к отдельным
участкам. Наиболее важным для зрительной
функции являются заднетеменная и
нижневисочная. Эти зоны осуществляют
сложную обработку зрительных сигналов,
связанную с построением образа и
переходом от описания отдельных признаков
объектов к описанию предметов и отношений
между ними. Восприятие сложных объектов,
описание их частей и соотношения между
ними проводится разными механизмами
мозга. По данным современных исследований,
эти механизмы локализованы в разных
областях коры. Механизм классификации
объектов и их инвариантное описание –
в нижневисочной коре, а механизм описания
пространственных отношений и полное
описание образа – в заднетеменной коре.
Эксперименты для подтверждения этих
механизмов и их локализации были
проведены на 3 группах обезьян. Первая
группа контрольная, у второй группы
билатерально удаляли нижневисочную
кору, а у животных билатеральных удаляли
заднетеменную кору. До проведения
операции у всех животных вырабатывали
пищевой двигательный условный рефлекс
на дифференцировку зрительно предъявляемой
фигуры, т.е. одну из 6 фигур, предъявляемых
одновременно, подкрепляли. Эту процедуру
проводили и после экстирпации коры.
После упрочения рефлекса проводили
преобразования фигур: меняли их размер
и ориентацию.
,
и дальше во многие другие части коры, в
том числе, теменной и височной. Таких
участков в коре насчитывают до 30 и они
связаны между собой волокнами в обоих
направлениях. Информация из стриарной
коры направляется в другие части мозга,
в т.ч. в базальные ядра и в промежуточный
мозг. Особенность: они связаны со
зрительной корой и относятся к отдельным
участкам. Наиболее важным для зрительной
функции являются заднетеменная и
нижневисочная. Эти зоны осуществляют
сложную обработку зрительных сигналов,
связанную с построением образа и
переходом от описания отдельных признаков
объектов к описанию предметов и отношений
между ними. Восприятие сложных объектов,
описание их частей и соотношения между
ними проводится разными механизмами
мозга. По данным современных исследований,
эти механизмы локализованы в разных
областях коры. Механизм классификации
объектов и их инвариантное описание –
в нижневисочной коре, а механизм описания
пространственных отношений и полное
описание образа – в заднетеменной коре.
Эксперименты для подтверждения этих
механизмов и их локализации были
проведены на 3 группах обезьян. Первая
группа контрольная, у второй группы
билатерально удаляли нижневисочную
кору, а у животных билатеральных удаляли
заднетеменную кору. До проведения
операции у всех животных вырабатывали
пищевой двигательный условный рефлекс
на дифференцировку зрительно предъявляемой
фигуры, т.е. одну из 6 фигур, предъявляемых
одновременно, подкрепляли. Эту процедуру
проводили и после экстирпации коры.
После упрочения рефлекса проводили
преобразования фигур: меняли их размер
и ориентацию.
Группа животных с удалённой нижневисочной корой при изменении размера фигуры на 25% резко снижало вероятность правильного опознания, а если размер фигуры уменьшался в 2 раза, то выбор становился случайным. Такие же результаты получили в этой группе при изменении ориентации фигур.
Животные с удалённой заднетеменной корой опознавали предмет при изменении размера и ориентации, причём опознавали даже лучше, чем контрольная группа. Т.е. разрушение задней теменной коры не нарушало оценки, которые инвариантно описывали объект. Нарушалось только восприятие пространственных отношений. Разрушение нижневисочной коры нарушало описание зрительного образа, и в этом случае исчезала целостность восприятия. Оказалось, что такой механизм присутствует только у высших животных, его нет у низших. Нейроны этой области не изменяют реакции на подкрепляемые и неподкрепляемые стимулы. В целом, исследование нейронов нижневисочной коры показало, что они могут быть разделены на три группы:
Первая группа реагирует на сенсорное значение стимулов и не соотносит его с моторными программами;
Вторая группа неоднородна, часть из этих нейронов реагирует на любое появление стимула в поле зрения, а другая часть – только при сопровождении стимула актом внимания;
Третья группа нейронов реагирует на сложные признаки образа, инвариантные преобразованиям изображения.
Этот механизм выделения признаков
нижневисочной корой был назван системой
«Что». Эта система служит для опознания
и категоризации объектов. Она начинается
от ганглиозных клеток сетчатки, волокна
которых поступают в наружное коленчатое
тело, затем в поле
![]() зрительной коры, далее в поле
зрительной коры, далее в поле
![]() ,
и затем в нижневисочную кору. Отдельные
признаки объекта обрабатываются
параллельно в различных зонах коры.
Локальное поражение одной из частей
коры нарушает восприятие одного признака
объекта. Например, ориентацию и длину
линий, сложные признаки предмета, цвет
и восприятие. Большая часть нейронов
задней теменной коры реагирует на
движение объектов в поле зрения и
управляет саккадами. Предполагают, что
в теменной коре локализованы механизмы,
которые служат для обработки сенсорной
информации и одновременно для генерации
действий. Поэтому заднетеменной коре
приписывают свойства создания модели
окружающего пространства, описание
локализаций и перемещение в этом
пространстве объектов по отношению к
телу наблюдателя. Кроме этого, область
заднетеменной коры определяет положение
и перемещение собственного тела в
пространстве. Таким образом, механизмы
заднетеменной коры соединяют два потока
информации: об окружающем пространстве
и о собственном положении наблюдателя
в этом пространстве. Эта система получила
название система «Где». Она начинается
от клеток сетчатки, волокна которых
поступают в верхние двухолмия. Верхние
слои клеток двухолмия реагируют на
зрительные образы, а глубокие слои
запускают саккадические движения глаз.
Амплитуда и направление саккад
обеспечивают попадание объекта в центр
поля зрения. Во время движения глаз
зрительные сигналы смещаются по сетчатке,
но образ зрительного мира не изменяется,
потому что рецептивные поля нейронов
двухолмия представлены участками
зрительного поля, а не участками сетчатки.
Зрительный образ дополняется сигналами
о движении глаз. Эта система формируется
после рождения в результате обучения,
что подтверждено экспериментами на
обезьянах.
,
и затем в нижневисочную кору. Отдельные
признаки объекта обрабатываются
параллельно в различных зонах коры.
Локальное поражение одной из частей
коры нарушает восприятие одного признака
объекта. Например, ориентацию и длину
линий, сложные признаки предмета, цвет
и восприятие. Большая часть нейронов
задней теменной коры реагирует на
движение объектов в поле зрения и
управляет саккадами. Предполагают, что
в теменной коре локализованы механизмы,
которые служат для обработки сенсорной
информации и одновременно для генерации
действий. Поэтому заднетеменной коре
приписывают свойства создания модели
окружающего пространства, описание
локализаций и перемещение в этом
пространстве объектов по отношению к
телу наблюдателя. Кроме этого, область
заднетеменной коры определяет положение
и перемещение собственного тела в
пространстве. Таким образом, механизмы
заднетеменной коры соединяют два потока
информации: об окружающем пространстве
и о собственном положении наблюдателя
в этом пространстве. Эта система получила
название система «Где». Она начинается
от клеток сетчатки, волокна которых
поступают в верхние двухолмия. Верхние
слои клеток двухолмия реагируют на
зрительные образы, а глубокие слои
запускают саккадические движения глаз.
Амплитуда и направление саккад
обеспечивают попадание объекта в центр
поля зрения. Во время движения глаз
зрительные сигналы смещаются по сетчатке,
но образ зрительного мира не изменяется,
потому что рецептивные поля нейронов
двухолмия представлены участками
зрительного поля, а не участками сетчатки.
Зрительный образ дополняется сигналами
о движении глаз. Эта система формируется
после рождения в результате обучения,
что подтверждено экспериментами на
обезьянах.
FILE 9