

Основные функции поверхностного аппарата
1. Трансмембранный транспорт. Представлен тремя категориями процессов:
а) диффузия;
б) пассивный транспорт или облегченная диффузия;
в) активный транспорт или избирательный (при участии АТФ и ферментов).
2. Рецепторная функция.
3. Транспорт в мембранной упаковке. Делится на экзоцитоз и эндоцитоз (фагоцитоз, пиноцитоз) в зависимости от того, в каком направлении переносятся вещества (в клетку или из клетки).
ЦИТОПЛАЗМАТИЧЕСКИЙ МАТРИКС. Представляет собой основную и наиболее важную часть клетки, ее истинную внутреннюю среду. Компоненты цитоплазматического матрикса осуществляют процессы биосинтеза в клетке и содержат ферменты, необходимые для продуцирования энергии, главным образом за счет анаэробного гликолиза.
Основные с в о и с т в а цитоплазматического матрикса:
1. С матриксом связаны коллоидные свойства клетки. Вместе с внутриклеточными мембранами вакуолярной системы его можно рассматривать как высокогетерогенную или многофазную к о л л о и д н у ю с и с т е м у.
2. Цитоплазматический матрикс обеспечивает изменение вязкости цитоплазмы различных клеток, которая возникает под действием внешних и внутренних факторов.
3. Цитоплазматический матрикс ответственен за циклоз, амебовидное движение, деление клетки и движение пигмента в хроматофорах.
4. Цитоплазматический матрикс определяет полярность расположения внутриклеточных компонентов.
5. Цитоплазматический матрикс обеспечивает механические свойства клеток, такие, как эластичность, способность к слиянию, ригидность.
ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ (ЭПС). ЭПС состоит из трех морфологических компонентов: канальцев, микровакуолей и крупных цистерн.
Существуют две разновидности ПС:
а) шероховатая или гранулярная, когда цистерны и канальцы связаны с рибосомами.
б) гладкая или агранулярная, когда связь с рибосомами отсутствует.
Функции ЭПС
1. Обеспечивает синтез белка.
.Пл
2. Обеспечивает активный транспорт различных соединений по внутримембранной фазе.
3. Синтез мембранных липидов.
4. Транспорт и накопление ионов в клетке, а также резервуар ионов кальция.
5. Синтез предшественников стероидных гормонов и других специфических соединений.
6. Детоксикация вредных продуктов метаболизма (особенно в гепатоцитах позвоночных).
7. Синтез протеолитических ферментов.
8. Первичный синтез секрета.
КОМПЛЕКС ГОЛЬДЖИ (КГ). КГ состоит из трех морфологических компонентов: уплощенные цистерны, микровакуоли и крупные цистерны (рис. 2).
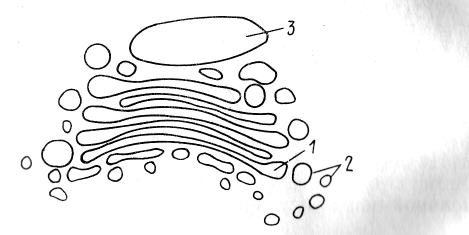
Рис. 2. Строение аппарата Гольджи: 1—уплощенные цистерны, 2 — микровакуоли, 3 — крупные цистерны
В растительных клетках цистерны отсутствуют. Такой КГ называют диктиосомы.
Особенность строения КГ
На своих мембранах КГ никогда не имеет рибосом.
Функции КГ
1. Формирование первичных лизосом с их своеобразными мембранами и сложной структурной организацией гидролаз.
2. Формирование особых структур — пероксисом или микротелец— пузырьков, ограниченных одинарной мембраной и содержащих каталазу, оксидазу Д-АМИНОКИСЛОТ, уротоксидазу и некоторые другие окислительные ферменты.
3. Сборка и «рост» мембран, которые затем окружают накапливающиеся продукты секреции, после чего они освобождаются из органеллы.
4. Обезвоживание, накопление, упаковка и транспорт продуктов секреции.
5. Синтез структурных компонентов клетки, таких, как, например, коллаген — компонент соединительной ткани.
6. Участвуют в синтезе желтка яйцеклеток и синтезе полисахароидов.
МИТОХОНДРИИ. Митохондрии ограничены двумя мембранами. Внешняя митохондриальная мембрана отделяет ее от гиалоплазмы. Обычно она имеет развитые контуры, не образует впячивания или складки. Внутренняя мембрана ограничивает собственно внутреннее содержимое митохондрий, ее матрикс. Характерной чертой внутренней мембраны митохондрий являются складки, которые имеют вид плоских гребней или крист. Как межмембранное пространство, так и полость матрикса заполнены содержимым гомогенного строения. В матриксе иногда выявляются тонкие нити и гранулы (рис. 6).
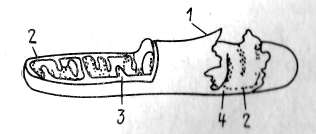
Рис.3. Строение митохондрии: 1 — наружная мембрана, 2— внутренняя мембрана, 3 — впячивания внутренней мембраны, 4 — места впячивания крист с поверхности внутренней мембраны
Функции митохондрий
1. Осуществляют синтез АТФ.
2. Участвуют в углеводном и азотистом обмене.
а). На наружной мембране и в окружающей ее гиалоплазме идут процессы анаэробного окисления (гликолиз).
б). На внутренней мембране митохондрий идут процессы, связанные с окислительным циклом трикарбоновых кислот и дыхательной цепью переноса электронов.
3. Осуществляют синтез белка. Митохондрии обладают полной системой синтеза белков, то есть имеют спою специфическую ДНК, митохондриальную ДНК
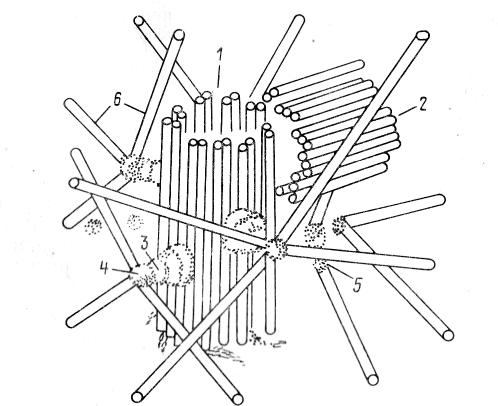
Рис. 4. Строение клеточного центра: 1—материнская центриоль, 2 — дочерние центриоли, 3 — ножка сателлита, 4 — головка сателлита, 5 — фокусы схождения микротрубочек, 6 — микротрубочки
КЛЕТОЧНЫЙ ЦЕНТР (диплосома) и центросферы носит название клеточного центра. Основу строения центриолей составляют расположенные по окружности девять групп микротрубочек (по три микротрубочки в группе), образующих таким образом полый цилиндр. Система микротрубочек центриоли описывается формулой (9+0), подчеркивая отсутствие микротрубочек в ее центральной части. Вокруг каждой центриоли расположен бесструктурный или тонковолокнистый матрикс(рис. 4).
Центриоли характерны и обязательны для клеток животных, их нет у высших растений, у низших грибов и некоторых простейших.
Строение и активность центриолей меняется в зависимости от периода клеточного цикла (интерфаза, митоз). Начиная с профазы и кончая телофазой, центриоли имеют сходное строение. В это время происходит формирование веретена митотического аппарата, состоящего из микротрубочек. То есть клеточный центр является местом полимеризации микротрубочек (рис. 8).
В предсинтетический период активность клеточного центра связана с образованием цитоплазматических микротрубочек (цитоскелета).
В синтетический период, помимо образования микротрубочек цитоскелета, может идти образование ресничек или жгутиков. Базальные тельца этих структур кроме периферических групп микротрубочек имеют центральную группу микротрубочек. Система микротрубочек реснички описывают как (9+1) в отличие от (9+0) системы центриолей.
При наступлении синтетического периода происходит удвоение числа центриолей. В постсинтетический период микротрубочки цитоскелета исчезают и вокруг центриолей (дипло-сом) начинают отрастать митотические трубочки.
Таким образом, функции клеточного центра заключаются:
а) в образовании цитоплазматических микротрубочек;
б) в построении веретена деления;
в) в образовании ресничек и жгутиков.
РИБОСОМЫ. Это сложные рибонуклеопротеиды (РИМ), и состав которых входят белки и молекулы РНК, примерно и равных весовых соотношениях. Работающая рибосома состоит из двух неравных субъединиц: большой и малой.

Рис. 5. Центриолярный цикл: а— диплосома во время митоза, б —центриоль
в начале G, — периода, в —центриоль в G, — периоде, г — центриоли в
S — периоде, удвоение центриолей, д — центриоли в G2 — периоде
субъединиц весьма сложна. Малая субъединица изогнута в виде телефонной трубки, а большая напоминает ковш.
Рибосомы присутствуют и в прокариотах, и в эукариотных клетках. В клетках эукариот существует две разновидности рибосом: рибосомы собственно цитоплазмы и рибосомы, локализованные в таких клеточных органоидах, как митохондрии и хлоропласты.
Рибосомы эукариот имеют коэффициент седиментации 80S коэффициент седиментации характеризует скорость осаждения частицы при ультрацентрифугировании), а прокариот — 70S. Рибосомы митохондрий и хлоропластов в общем тоже относятся к классу 70S.
В состав цитоплазматических рибосом эукариотных клеток входят всего четыре молекулы РНК (из них три в большой субъединице), а в рибосомах прокариот присутствуют три молекулы РНК (из них две в большой субъединице).
Белковый состав рибосом очень неоднороден. Белки большой и малой субъединиц рибосом резко отличаются по аминокислотному составу и молекулярной массе.
При укладке тяжа РНП в СУБЪЕДИНИЦЫ рибосом образуются белковые активные центры. На малой субъединице имеются два активных центра:
а) и-РНК — связывающий участок;
б) участок, удерживающий амнноацил-т-РНК. На большой субъединице различают:
а) аминоацильный центр;
б) пептидный центр.
Между этими участками выделяют еще один, частично перекрывающийся с ним, - дипептидилтрансферазный центр, который катализирует образование пептидных связей.
Рибосомы — это фабрики биосинтеза.
ЛИЗОСОМЫ. Состоят из очень пестрого класса пузырьков размером 0,2—0,8 мкм, ограниченных одиночной мембраной. Каждая лизосома содержит разнообразные ферменты, способные расщеплять биологические продукты в слабокислой среде. Содержащиеся в лизосоме ферменты окружены мембраной и благодаря этому разобщены с субстратом, находящимся в клетке.
Основные протеолитические ферменты лизосом:
1. Кислая РНКаза и кислая ДНКаза расщепляют нуклеиновые кислоты.
2. Кислая фосфотаза — фосфорные эфиры.
3. Фосфопротеидфосфотаза — фосфопротеиды и пирофосфа-ты.
4. Катепсин и коллагеназа — белки.
5. Альфа-глюкозидаза — полисахариды.
6. Бетта-глюкоронидаза — мукополисахариды.
7. Арилсульфатаза — эфиры серной кислоты.
Среди различных по морфологии лизосомных частиц можно выделить по крайней мере четыре типа:
1. Первичные лизосомы формируются из вакуолей периферической зоны аппарата Гольджи и заполнены протеолитическими ферментами в неактивной форме, ранее синтезированными на гранулярной эндоплазматической сети (рис. 9).
2. Вторичные лизосомы или внутриклеточные пищеварительные вакуоли по своей морфологии представляют собой первичную лизосому, слившуюся с фагоцитарными или пиноцитозными вакуолями. От типа поглощенных веществ или частичек зависит размер и внутренняя структура таких лизосом. Судьба поглощенных биогенных веществ, попавших и состав лизосомы, заключается в их расщеплении гидролазами до мономеров и в транспорте этих мономеров через мембрану лизосомы в состав цитоплазмы, где они включаются в различные синтетические и обменные процессы.
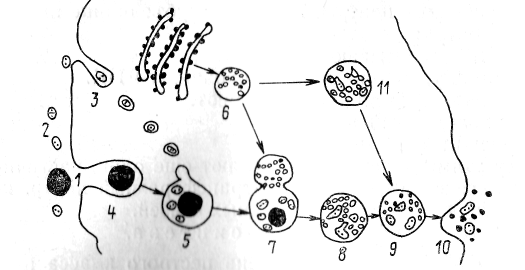
Рис. 6. 1,12 —крупные и мелкие частицы в среде, окружающей клетку, 3,4 — захват частиц путем фагоцитоза и возникновение лизосом (первичная лизосома), 7 — фаголизосома, 8 —пищеварительное тельце, 9 — остаточное тельце, 10 — выделение продуктом разложения 11 —аутолизосомы
3. Телолизосомы или остаточные тельца. Такие лизосомы возникают на вторичных, при переваривание биогенных микромолекул внутри лизосом идет не до конца. Остаточные тельца уже содержат много протеолитических ферментов, в них происходит уплотнение содержимого и его отложение в виде сложных структур. Судьба остаточных телец может быть двоякой: одни из них выбрасываются из клетки путем экзоцитоза, другие же остаются в клетках вплоть до их гибели (например, липофусциновые гранулы).
4. Аутолизосомы (аутофагосомы). По своей морфологии их относят к вторичным лизосомам, по г тем отличием, что в составе этих вакуолей встречаются фрагменты или даже целые цитоплазматические структуры. В этом случае лизосомы выполняют роль внутриклеточных чистильщиков, контролирующих дефектные структуры.