

Вегетативная иннервация отдельных органов (частная вегетология)
На основании изложенных выше общих закономерностей строения и функционального назначения вегетативного отдела единой нервной системы необходимо более подробно рассмотреть и систематизировать данные, касающиеся иннервации и некоторых механизмов регуляции функций отдельных органов. При этом нужно иметь в виду, что специфика вегетативной иннервации каждого органа определяется его морфофункциональными особенностями, в частности зависит от наличия и строения гладкомышечных компонентов, железистого аппарата, органоспецифичности сосудистого русла, режимов функциональной деятельности. Кроме того, не следует забывать об универсальности распространения (во всяком случае – симпатического отдела) и адаптационно-трофической роли вегетативной нервной системы.
ИННЕРВАЦИЯ ГЛАЗНОГО ЯБЛОКА
Гладкомышечная ткань глазного яблока представлена двумя мышцами радужки – мышцей, суживающей зрачок, m. sphincter pupillae, и мышцей, расширяющей зрачок, m. dilatator pupillae, а также мышцей ресничного тела, m. ciliаris, состоящей из меридионально, циркулярно и радиально ориентированных мышечных пучков. Действие циркулярно расположенных и радиальных мышц радужки обеспечивает соответственно сужение или расширение зрачка (зрачковый рефлекс); комплексное напряжение переплетающихся слоев ресничной мышцы ведет к расслаблению ресничного пояска (цинновой связки), посредством которого хрусталик фиксируется по экватору к ресничному телу. В результате этого, в силу расслабления капсулы хрусталика и эластичности хрусталиковых волокон, кривизна и преломляющаяся способность его увеличиваются, глаз устанавливается на рассматривание близлежащих предметов. При расслаблении ресничной мышцы цинновы связки натягиваются (обратный эффект), хрусталик приобретает плоскую форму, глаз устанавливается на рассматривание удаленных предметов. Способность глаза к ясному видению различно расположенных предметов носит название аккомодации глаза.
В обеспечении зрачкового рефлекса принимают участие как симпатический, так и парасимпатический отделы вегетативной нервной системы. При этом симпатический отдел иннервирует мышцу, расширяющую зрачок, а парасимпатический – сфинктер зрачка и мышцу ресничного тела, осуществляя сужение зрачка и аккомодацию глаза.
Спинномозговые центры симпатической иннервации органов головы, в том числе и мышцы, расширяющей зрачок, располагаются в латеральном промежуточном ядре боковых рогов серого вещества спинного мозга на уровне CVIII, ThI-ThII сегментов. Преганглионарные волокна от клеток этого ядра выходят в составе передних корешков и соответствующих указанным сегментам спинномозговых нервов и в виде белых соединительных ветвей достигают верхних грудных узлов симпатического ствола. Не прерываясь в них, они транзитом восходят по межузловым ветвям симпатического ствола до первого шейного узла, где переключаются на последний эфферентный нейрон.
Верхний шейный узел, ganglion cervicalis superius, является самым большим узлом в системе симпатического ствола (до 2 см длины). Он располагается спереди поперечных отростков II-III шейных позвонков на длинной мышце головы, покрытой предпозвоночной пластинкой собственной фасции шеи. Наиболее крупная ветвь верхнего шейного узла – внутренний сонный нерв, n. caroticus internus. По ходу одноименной артерии он образует plexus caroticus internus, и, пройдя вместе с артерией через сонный канал, распределяется по ветвям внутренней сонной артерии. По ходу глазной артерии одноименное симпатическое сплетение через канал зрительного нерва достигает глазницы. Здесь постганглионарные симпатические волокна в составе длинных (от носоресничного нерва I ветви тройничного нерва) и коротких ресничных нервов (от ресничного узла) поступают в глазное яблоко для иннервации мышцы, расширяющей зрачок, и сосудов глазного яблока (сосудосуживающий эффект).
Сложные двухнейронные эфферентные пути симпатической иннервации содержимого глазницы обусловливают соответствующие симптомокомплексы и своеобразие клинических проявлений при нарушении проводимости в них. Так, при повреждении спинного мозга на уровне CVIII, ThI-ThII сегментов, наряду с двигательными и (или) чувствительными соматическими расстройствами, возникают, казалось бы, мало связанные с местом поражения симптомы, известные в неврологии под названием синдрома Хорнера – Клод Бернара: сужение зрачка - за счет преобладания парасимпатической иннервации и сокращения сфинктера радужки, уменьшение глазной щели, западание глазного яблока. Поэтому клиницисты группу клеток, заложенных в боковых рогах спинного мозга на уровне CVIII, ThI-ThII сегментов, трактуют как centrum ciliarespinalis. Аналогичные симптомы проявляются и при изолированном поражении верхнего шейного узла симпатического ствола, откуда исходит внутренний сонный нерв, иннервирующий большинство органов и сосудов головы.
Парасимпатическая иннервация гладкомышечных элементов и сосудов глаза осуществляется за счет добавочного ядра глазодвигательного нерва (III пара черепных нервов), nucleus oculomotorius accessoriиs (парасимпатическое ядро Якубовича, Вестфаль - Эдингера). Как было отмечено выше, это ядро располагается на дне водопровода среднего мозга, на уровне верхнего двухолмия и вентральнее моторного ядра III пары. Прегаглионарные волокна от клеток добавочного ядра проходят в составе глазодвигательного нерва до глазницы. На дне ее, латерально от зрительного нерва, располагается ресничный узел, ganglion ciliare, на клетках которого синаптически заканчиваются преганглионарные волокна, исходящие из добавочного ядра глазодвигательного нерва. Постганглионарные волокна от клеток ресничного узла в составе коротких ресничных нервов, nn. ciliares breves, входят в глазное яблоко и иннервируют мышцу, суживающую зрачок, и ресничную мышцу.
Роль афферентного звена в системе рефлекторной дуги зрачкового рефлекса и аккомодации глаза выполняют проводящие пути зрительного анализатора. Как известно, оптические раздражения от сетчатки глаза по зрительным нервам (II пара), а после частичного перекреста их (chiasma opticum) – по зрительным трактам (tractus opticus) поступают в подкорковые центры зрения: латеральные коленчатые тела, corpori geniculati lateralеs, подушку, pulvinar, зрительных бугров и верхние холмики, colliculi superiorеs, крыши среднего мозга (tectum mesencephali). Клетки серого вещества верхнего двухолмия являются ассоциативными нейронами, которые связывают орган зрения со многими ядрами черепных нервов, в том числе с ядрами глазодвигательных нервов и парасимпатическим ядром Якубовича. Причем связь с последним – двусторонняя, поэтому при попадании света на один глаз суживается зрачок и другого глаза. Кроме того, верхнее двухолмие связано с ядрами передних столбов (рогов) спинного мозга (tractus tectospinalis, fasciculus longitudinalis medialis – от ядер Кахаля и Даркшевича). Благодаря этому достигаются сочетанные функции мышц глазного яблока и шеи (сочетанный поворот головы и глаз в одну сторону) и защитные двигательные реакции в ответ на световые раздражения.
По современным представлениям (Гайворонский И.В., 2000), связь надсегментарных центров вегетативной регуляции с сегментарными центрами (центральными эффекторными нейронами) симпатического и парасимпатического отделов, заложенными в стволовой части головного мозга, в тораколюмбальном и сакральном отделах спинного мозга, осуществляется посредством заднего продольного пучка, fasciculus longitudinalis dorsalis (пучок Шютца). Он начинается от ядер гипоталамуса, как основного надсегментарного центра «вегетативной жизни», проходит через ствол головного мозга, где отдает коллатерали к парасимпатическому ядру Якубовича (средний мозг), верхнему слюноотделительному ядру (мост мозга), нижнему слюноотделительному ядру и дорсальному ядру блуждающего нерва (продолговатый мозг). Далее этот пучок спускается в боковых канатиках спинного мозга, примыкая к переднему краю бокового пирамидного тракта, и посегментно заканчивается на симпатических центрах латерального промежуточного ядра (торако-люмбальный отдел) и парасимпатических центрах сакрального отдела спинного мозга (SII-SIV). Таким образом зрительный анализатор замыкается соответственно на парасимпатическом ядре глазодвигательного нерва и реснично-спинальном симпатическом центре (CVIII, ThI-ThII), благодаря чему осуществляется координация и динамизм зрачкового рефлекса.
ИННЕРВАЦИЯ СЛЕЗНОЙ ЖЕЛЕЗЫ
Секреторная деятельность слезной железы находится под постоянным контролем со стороны как симпатического, так и парасимпатического отделов вегетативной нервной системы.
Симпатические сегментарные центры для иннервации слезной железы располагаются в латеральном промежуточном ядре, nucleus intermediolateralis, верхних грудных сегментов спинного мозга (ThI-ThII). От нейронов этого ядра преганглионарные волокна следуют по уже описанным выше путям (передние корешки спинного мозга, верхние грудные спинномозговые нервы, белые соединительные ветви, межузловые ветви) и достигают верхнего шейного симпатического узла, где синаптически переключают нервный импульс на последний эффекторный нейрон.
Постганглионарные волокна от верхнего шейного узла идут в составе внутреннего сонного нерва и соименного сплетения, затем продолжаются на сплетения глазной и слезной артерий, достигая слезной железы. Симпатический отдел вегетативной нервной системы уменьшает либо тормозит секрецию слезной жидкости. Поэтому общая картина повышения тонуса «симпатикуса» характеризуется блестящими, выпуклыми глазами, расширенными зрачками, сухостью конъюнктивального мешка (Триумфов А.В., 1964).
Парасимпатическая иннервация слезной железы осуществляется за счет верхнего слюноотделительного ядра, nucleus salivatorius superior, находящегося в мосту мозга (рис. 15). Некоторые авторы выделяют в нем специальную группу нейронов под названием слезного ядра, nucleus lacrimalis (Гайворонский И.В., 2000). Тем не менее, все преганглионарные волокна проходят в составе лицевого нерва (его промежуточной порции), в пирамиде височной кости они отделяются, образуя большой каменистый нерв, n. petrosus major, и входят в состав барабанной струны, chorda thympani. Большой каменистый нерв через свое отверстие (hiatus) выходит на переднюю поверхность пирамиды и в составе нерва крыловидного канала, n. canalis pterygoidei (совместно с симпатическими постганглонарными волокнами, n. petrosus profundus), проходит в крыловидно-небную ямку. Здесь находится крылонебный парасимпатический узел, ganglion pterygopalatinum, на клетках которого и заканчиваются преганглионарные волокна большого каменистого нерва.
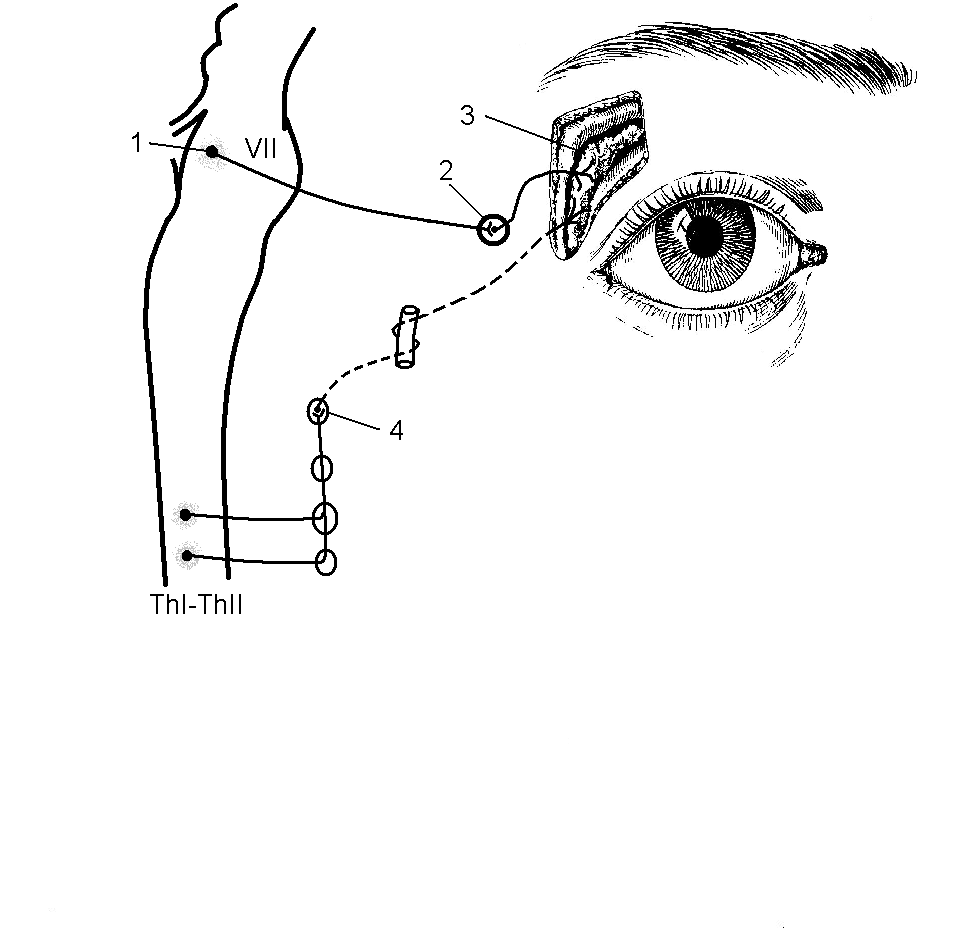
Рис. 15. Иннервация слезной железы:
1 – nucleus salivatorius superior; 2 – ganglion pterygopalatinum; 3 – glandula lacrimalis; 4 – ganglion cervicale superius;
VII – n. facialis; ThI–ThII – сегменты спинного мозга.
Постганглионарные волокна от клеток крылонебного узла присоединяются вначале к верхнечелюстному нерву (II ветвь тройничного нерва), затем – к скуловому нерву и в составе слезного нерва (соматически-чувствительного) достигают слезной железы. Парасимпатические нервные структуры (верхнее слюноотделительное ядро, крылонебной узел) увеличивают секрецию слезной жидкости. При поражении этих образований или проводящих путей и при преобладании симпатического тонуса происходит уменьшение слезоотделения, наблюдается сухость конъюнктивального мешка и роговицы глаза.
Вполне естественно, что симпатические и парасимпатические сегментарные центры, регулирующие секрецию слезной жидкости (здесь проявляется синергизм деятельности обоих отделов), находятся под постоянным влиянием и контролем со стороны вышестоящих надсегментарных центров. Так, экспериментальные и клинические наблюдения показывают, что при поражении премоторной зоны лобной доли, при раздражении полосатого тела (хвостатого и чечевицеобразного ядер) возникает обильное слезотечение. С ядрами гипоталамуса верхнее слюноотделительное ядро и латеральное промежуточное ядро серого вещества спинного мозга связывает упомянутый выше задний продольный пучок. Благодаря этому достигаются согласованность и четкая координация деятельности слезной железы.
ИННЕРВАЦИЯ СЛЮННЫХ ЖЕЛЕЗ
Постоянная выработка, качество и количество слюны, поступающей в полость рта, регулируются и контролируются обоими звеньями вегетативной нервной системы: большие и малые слюнные железы получают как симпатическую, так и парасимпатическую иннервацию. Афферентация желез осуществляется за счет чувствительных ветвей тройничного нерва.
Центры симпатической иннервации всех слюнных желез (околоушной, поднижнечелюстной, подъязычной, малых желез слизистой оболочки полости рта) располагаются в латеральном промежуточном ядре серого вещества спинного мозга на уровне верхних грудных сегментов. Преганглионарные волокна от нейронов nucleus intermediolateralis следуют в составе вначале передних корешков спинного мозга, а затем в составе соответствующих спинномозговых нервов. Отделившись от них, волокна в виде белых соединительных ветвей подходят к грудному отделу симпатического ствола и по межузловым ветвям достигают верхнего шейного узла. Здесь происходит синаптическое переключение преганглионарных волокон на последний эфферентный нейрон рефлекторной дуги (рис.16).
От верхнего шейного узла отходят 2-3 стволика постганглионарных волокон, которые образуют вокруг наружной сонной артерии одноименное нервное сплетение, plexus caroticus externus. Последнее внизу связано с общим сонным сплетением, plexus caroticus communis, - от среднего шейного узла, а при его отсутствии – от нижнего (или звездчатого) шейного узла. Наружное сонное сплетение по ветвям артерии переходит в соответствующие периартериальные нервные сплетения, в частности: по лицевой артерии оно достигает поднижнечелюстной слюнной железы и малых губных желез, по язычной артерии – подъязычной слюнной железы и язычных желез, по поверхностной височной артерии – околоушной слюнной железы, по верхнечелюстной артерии – малых слюнных желез слизистой оболочки неба и щеки.
Симпатический отдел вегетативной нервной системы вызывает уменьшение секреции слюны, повышает вязкость ее. Малообильная густая слюна обусловливает относительную сухость слизистой оболочки полости рта, что характерно, например, для аффекторов страха, гнева и т.п.
Парасимпатическая
иннервация больших и малых слюнных
желез отличается более специализированной
структурой своих центров и топографическим
разнообразием эфферентных проводящих
путей. Специфическим центром
парасимпатической иннервации
поднижнечелюстной и подъязычной слюнных
желез и большинства малых желез слизистой
оболочки полости рта является верхнее
слюноотделительное ядро, nucleus
salivatorius
superior,
расположенное в ромбовидной ямке в
области моста (см. рис. 16). Преганглионарные
волокна от нейронов этого узла выходят
из моста вместе с соматически двигательными
волокнами лицевого нерва (VII
пара). В канале лицевого нерва пирамиды
височной кости преганглионарные
парасимпатические волокна отделяются
от этого нерва и расходятся в двух
направлениях. Часть волокон образуют
большой
каменистый нерв,
n.
petrosus
majoris,
который в составе нерва крыловидного
канала проникает в крыловидно-н ебную
ямку и заканчивается на клетках
крылонебного
узла,
ganglion
ебную
ямку и заканчивается на клетках
крылонебного
узла,
ganglion
pterygopalatinum.
Рис. 16. Вегетативная иннервация слюнных желез:
1 – nucleus salivatorius superior; 2 - nucleus salivatorius inferior; 3 – ganglion cervicale superius; 4 – glandula submandibularis; 5 – glandula sublingualis; 6 – glandula parotidea; 7 – ganglion submandibulare; 8 – ganglion sublinguale; 9 – ganglion oticum; VII – n. facialis; IX – glossopharyngeus; ThI-ThII – сегменты спинного мозга.
Постганглионарные волокна эфферентных нейронов этого узла присоединяются к чувствительным ветвям верхнечелюстного нерва (II ветвь тройничного нерва) для иннервации желез слизистой оболочки полости носа (задние верхние и нижние носовые ветви, rr. nasals posteriorеs superiorеs et inferiorеs) и малых небных слюнных желез (большой и малый небные нервы, nn. palatine majores et minores, носонебный нерв, n. nasopalatinus).
Другая часть преганглионарных парасимпатических волокон, первоначально входящих в состав лицевого нерва, совместно с периферическими отростками псевдоуниполярных клеток чувствительного узла промежуточного нерва – узла коленца, ganglion geniculi, образуют барабанную струну, chorda thympani. Она выходит из пирамиды височной кости через каменисто-барабанную щель, fissura petrothympanica, и между латеральной и медиальной крыловидными мышцами присоединяется к язычному нерву, n. lingualis (ветвь нижнечелюстного нерва, V пара). В составе этого нерва преганглионарные парасимпатические волокна подходят к поднижнечелюстному и подъязычному узлам, ganglion submandibulare et ganglion sublinguale.
Ganglion submandibulare расположен рядом с язычным нервом на медиальной поверхности одноименной слюнной железы. Сравнительно короткие постганглионарные волокна от клеток узла, вместе с симпатическими волокнами от периартериального сплетения язычной артерии, проникают в поднижнечелюстную слюнную железу. Ganglion sublinguale лежит на наружной поверхности подъязычной слюнной железы. Постганглионарные волокна от него обеспечивают парасимпатическую иннервацию подъязычной, малых губных и язычных желез.
Парасимпатическая иннервация околоушной слюнной железы, glandula parotis, осуществляется за счет нижнего слюноотделительного ядра, nucleus salivatorius inferior, расположенного в продолговатом мозге (см. рис. 16). Преганглионарные волокна от нейронов этого ядра входят в состав языкоглоточного нерва, n. glossopharyngeus (IX пара черепных нервов). По выходе нерва из яремного отверстия от него отходит барабанный нерв, n. tympanicus, состоящий из преганглионарных парасимпатических волокон - от нижнего слюноотделительного ядра и чувствительных волокон – от нижнего узла языкоглоточного нерва. Барабанный нерв через одноименный каналец проходит в барабанную полость, образуя в слизистой оболочке ее барабанное сплетение. В его состав входят также симпатические волокна от сонно-барабанных нервов из внутреннего сонного сплетения. Таким образом, смешанный состав барабанного сплетения обеспечивает, с одной стороны, чувствительную иннервацию слизистой оболочки барабанной полости и слуховой трубы (среднее ухо), а с другой, - вегетативную, симпатическую и парасимпатическую иннервацию ее железистого аппарата.
Оставшаяся часть преганглионарных парасимпатических волокон барабанного нерва (сплетения) под названием малого каменистого нерва, n. petrosus minor, выходит на переднюю поверхность пирамиды височной кости и через рваное или специальное отверстие в больших крыльях клиновидной кости (канал Арнольда) достигает ушного узла, ganglion oticum. Последний располагается на наружном основании черепа медиально от нижнечелюстного нерва (III ветвь тройничного нерва) и овального отверстия, через которое этот нерв выходит из полости черепа. Постганглионарные ветви присоединяются к проходящему здесь ушно-височному нерву, n. auriculotemporalis, и вступают в околоушную железу.
Парасимпатический отдел вегетативной нервной системы усиливает секреторную функцию слюнных и слизистых желез, в частности обусловливает выработку и выделение обильной жидкой слюны белкового характера. Однако, постоянная смена режима секреции, разные количество и состав слюны возможно только при морфофункциональной связи симпатических и парасимпатических центров с вышестоящими вегетативными центрами, куда поступают интероцептивные сигналы. Подтверждением тому могут служить наиболее очевидные клинические признаки обильного слезо- и слюнотечения при поражении лобных долей большого мозга, при раздражении базальных ядер и ядер гипоталамуса и других надсегментарных центров. Общим интегрирующим путем, связывающим надсегментарные и сегментарные симпатические и парасимпатические центры, считается задний продольный пучок, fasciculus longitudinalis dorsalis. Важная роль в этом отводится ретикулярной формации и ее нисходящим проводящим путям (Хауликэ И., 1978).
ИННЕРВАЦИЯ ОРГАНОВ ШЕИ
Непосредственными тканевыми объектами вегетативной иннервации органов шеи являются секреторный (железистый) аппарат слизистой оболочки и сосуды глотки, гортани, начальных отделов пищевода и трахеи, а также щитовидная и паращитовидная железы. Все они имеют двойную иннервацию – симпатическую и парасимпатическую. Кроме того, в образовании нервных сплетений этих органов принимают участие соматически чувствительные волокна спинномозговых, языкоглоточного и блуждающего нервов.
Полые органы. Симпатическая иннервация глотки, гортани, шейного отдела пищевода осуществляется, в основном, за счет наружного сонного сплетения, plexus caroticus externus, являющегося производным верхнего шейного узла симпатического ствола. Как уже отмечалось, центры симпатической эфферентации органов головы и шеи находятся в латеральном промежуточном ядре (веществе) верхних грудных сегментов спинного мозга (ThI-ThII). Постганглионарные волокна от нейронов верхнего шейного узла продолжаются на ветви наружной сонной артерии и достигают рабочих органов. Кроме того, в симпатической иннервации полых органов шеи принимают участие гортанно-глоточные ветви, rr. laryngopharyngei, непосредственно отходящие от верхнего шейного симпатического узла. Они входят в состав глоточного, пищеводного и гортанного сплетений.
Сообразно общей функциональной роли и адренергической медиации нейро-железистых синапсов, симпатический отдел вегетативной нервной системы уменьшает секреторную функцию желез. Поэтому при повышении тонуса мозговых или периферических (узловых) симпатических центров возникает сухость не только во рту, но и в начальных отделах пищеварительного и дыхательного трактов.
Парасимпатическая иннервация полых органов шеи осуществляется за счет ветвей блуждающего нерва, n. vagus (X пара). Сегментарным парасимпатическим центром для него является дорсальное ядро, nucleus dorsalis n. vagi, расположенное в нижнем углу ромбовидной ямки в продолговатом мозге. В шейном отделе от блуждающего нерва отходят преганглионарные парасимпатические волокна в составе смешанных глоточных ветвей, rr. pharyngei, верхнего гортанного нерва, n. laryngeus superior, и конечной ветви возвратного гортанного нерва, n. laryngeus recurrens, - нижнего гортанного нерва. Все они принимают участие в образовании сложных подслизистых нервных сплетений, состоящих из чувствительных ветвей языкоглоточного и блуждающего нервов, преганглионарных парасимпатических волокон блуждающего нерва, из постганглионарных симпатических волокон соответствующих периартериальных сплетений и гортанно-глоточного нерва. В интрамуральных (внутристеночных) нервных сплетениях находятся нервные клетки, на которых заканчиваются преганглионарные парасимпатические волокна блуждающего нерва. Постганглионарные волокна ганглиозно-сетчатых сплетений подходят к концевым отделам желез, протокам и сосудам. Парасимпатический отдел вегетативной нервной системы усиливает секреторную деятельность желез слизистой оболочки полых органов шеи.
Щитовидная и паращитовидные железы. Источниками симпатической иннервации для щитовидной и паращитовидных желез являются общее сонное сплетение, plexus caroticus communis, и подключичное сплетение, plexus subclavius. Общее сонное сплетение формируется из постганглионарных волокон среднего шейного симпатического узла, а при его отсутствии – из нижнего шейного или звездчатого (шейно-грудного) узла. Преганглионарные волокна к этим узлам подходят от латерального промежуточного ядра верхних грудных сегментов спинного мозга. Подключичное нервное сплетение (вокруг одноименной артерии) образуется ветвями шейно-грудного симпатического узла.
Постганглионарные симпатические волокна из общего сонного и подключичного сплетений продолжаются на ветви соименных артерий, в частности в виде периартериальных сплетений по ходу верхней и нижней щитовидных артерий подходят к секреторному аппарату щитовидной и паращитовидных желез. Имея адренергическую медиацию нейротканевых синапсов, симпатические волокна оказывают тормозящее влияние на секреторную функцию этих желез.
Парасимпатическая иннервация щитовидной и паращитовидных желез обеспечивается дорсальным ядром блуждающего нерва. Часть преганглионарных волокон его, проходящих в составе возвратного гортанного нерва (ветвь X пары), в области шеи отделяется в виде ветвей щитовидной железы, rr. glandulae thyroideae, к щитовидной и паращитовидным железам. Синаптический перерыв преганглионарных волокон, видимо, происходит на клетках ганглиозно-сетчатых образований желез, однако морфофизиологические основы такой медиации недостаточно выяснены. В связи с этим следует отметить, что такой же «физиологической загадкой» пока остается морфологический факт наличия нервных клеток по ходу блуждающего нерва. Возможно, что внутриствольные нейроны являются своеобразными ганглионарными образованиями, постганлионарные волокна которых принимают участие в парасимпатической иннервации некоторых органов шеи.
Сегментарные симпатические и парасимпатические центры – «команды» находятся под постоянным влиянием со стороны высших центров вегетативной регуляции. Связь их с дорсальным ядром блуждающего нерва и торако-люмбальными симпатическими центрами спинного мозга осуществляется посредством заднего продольного пучка, который начинается от ядер гипоталамуса. Проходя через ствол мозга, он отдает коллатерали к nucleus dorsalis n. vagi, в области спинного мозга – к латеральному промежуточному ядру, благодаря чему осуществляется содружественная деятельность симпатических и парасимпатических центров.
Интегрирующая роль высших центров вегетативной нервной системы особо важна для гормональной деятельности щитовидной и паращитовидных желез, принимающих непосредственное участие в регуляции обмена веществ, процессов роста и размножения тканей. Именно на уровне этих центров происходит замыкание соматических и интерорецептивных рефлексов, в ответ на которые эндокринные железы меняют уровень своей деятельности и адаптируют трофические процессы в ответ на изменения внутренней и внешней среды.
ИННЕРВАЦИЯ ОРГАНОВ ДЫХАНИЯ
Вегетативная иннервация верхних дыхательных путей была рассмотрена в предыдущих разделах пособия. Здесь представлены морфофизиологические особенности иннервации трахеи, бронхов и легких.
Симпатическая иннервация трахеи и бронхов происходит за счет соименных ветвей, отходящих от звездчатого и верхних грудных узлов симпатического ствола и образующих соответственно plexus trachealis et plexus bronchialis. По ходу бронхов они продолжаются в легочное сплетение, plexus pulmonalis. Мозговыми центрами симпатической иннервации органов дыхания считаются участки латерального промежуточного ядра на протяжении от II до VI грудного сегмента спинного мозга (Турыгин В.В., 1988). Преганглионарные волокна от нейронов этого ядра подходят по обычным для симпатических волокон путям к грудным узлам (передние корешки, ThII-ThVI спинномозговые нервы, белые соединительные ветви). Постганглионарные волокна следуют от узлов либо самостоятельно, либо в составе грудного аортального сплетения и иннервируют гладкомышечные элементы, железы слизистой, сосуды трахеи и бронхов.
Повышение тонуса симпатических центров регуляции дыхания вызывает расширение бронхов, сужение просвета сосудов, уменьшение секреции желез. Поэтому картина так называемой «симпатикотонии» (например, при аффектах страха, испуга и других) характеризуется свободным учащенным дыханием, сухостью слизистых и т.п. Такое состояние обычно сочетается с увеличением силы и частоты сердечных сокращений (тахикардия), что при расширенных бронхах и учащенном дыхании соответствует повышению газообмена в легких.
Парасимпатическая иннервация органов дыхания осуществляется за счет дорсального ядра блуждающего нерва. Преганглионарные волокна его в составе возвратного гортанного нерва, n. laryngeus recurrens (ветвь X нерва), подходят к шейному отделу трахеи и прерываются на клетках интрамурального нервного сплетения. Короткие постганглионарные волокна от ганглиозных клеток иннервируют гладкую мускулатуру и железы слизистой оболочки трахеи.
От грудных отделов блуждающих нервов отходят трахеальные и бронхиальные ветви, которые по ходу бронхиального дерева и сосудов продолжаются в легочное сплетение. Последнее представлено чувствительными (от нижнего узла X пары), преганглионарными парасимпатическими волокнами (от заднего ядра X нерва) и постганглионарными симпатическими волокнами (от грудных узлов симпатического ствола). Преганглионарные парасимпатические волокна вступают в контакт с нервными клетками ганглиозных скоплений легких или клетками, расположенными по ходу бронхов и сосудов. Короткие аксоны этих клеток подходят к гладкомышечным структурам бронхов и железам слизистой оболочки. Парасимпатические волокна вызывают сужение бронхов, усиление секреции слизистой жидкости.
ИННЕРВАЦИЯ СЕРДЦА
Сердце отличается значительным количеством источников симпатической и парасимпатической иннервации, большой сложностью строения экстра- и интракардиальных нервных сплетений, наличием в них различных скоплений нервных клеток и ганглиев. Все это обусловлено спецификой строения и функции миокарда, необходимостью четкой последовательности и регламентированности сокращений предсердий и желудочков, постоянно меняющимися силой и частотой напряжения сердечной мышцы.
Спинномозговые центры симпатической иннервации сердца находятся в боковом промежуточном ядре на протяжении пяти-шести верхних грудных сегментов спинного мозга. Преганглионарные волокна от нейронов этого ядра обычными для них путями (передние корешки, ThI-ThV спинномозговые нервы, белые соединительные ветви) достигают симпатических стволов и синаптически вступают в связь с клетками верхних грудных и трех шейных узлов. От каждого шейного узла отходят постганглионарные ветви, образующие соответственно верхний, средний и нижний шейные сердечные нервы, nn. cardiaci cervicales superior, medius et inferior. Постганглионарные волокна от грудных узлов (ThII-ThV) составляют грудные сердечные нервы, nn. cardiaci thoracici. Существует мнение, что часть преганглионарных волокон проходит транзитом через симпатический ствол и заканчивается на нервных клетках сердечных сплетений (рис. 17).
С физиологических и клинических позиций несомненный интерес представляют топография и распределение шейных сердечных нервов. Так, правые верхний и средний сердечные нервы спускаются вдоль плечеголовного ствола и входят в состав глубокого сердечного сплетения, расположенного между дугой аорты и трахеей у места ее бифуркации. Левые верхний и средний шейные сердечные нервы проходят вдоль левой общей сонной артерии. В области аорты левый средний нерв продолжается в глубокое сердечное сплетение, а левый верхний нерв входит в состав поверхностного сердечного сплетения, которое находится между (под) дугой аорты и легочным стволом у места его раздвоения. Нижние шейные сердечные нервы, как правило, переходят в глубокое сердечное сплетение.
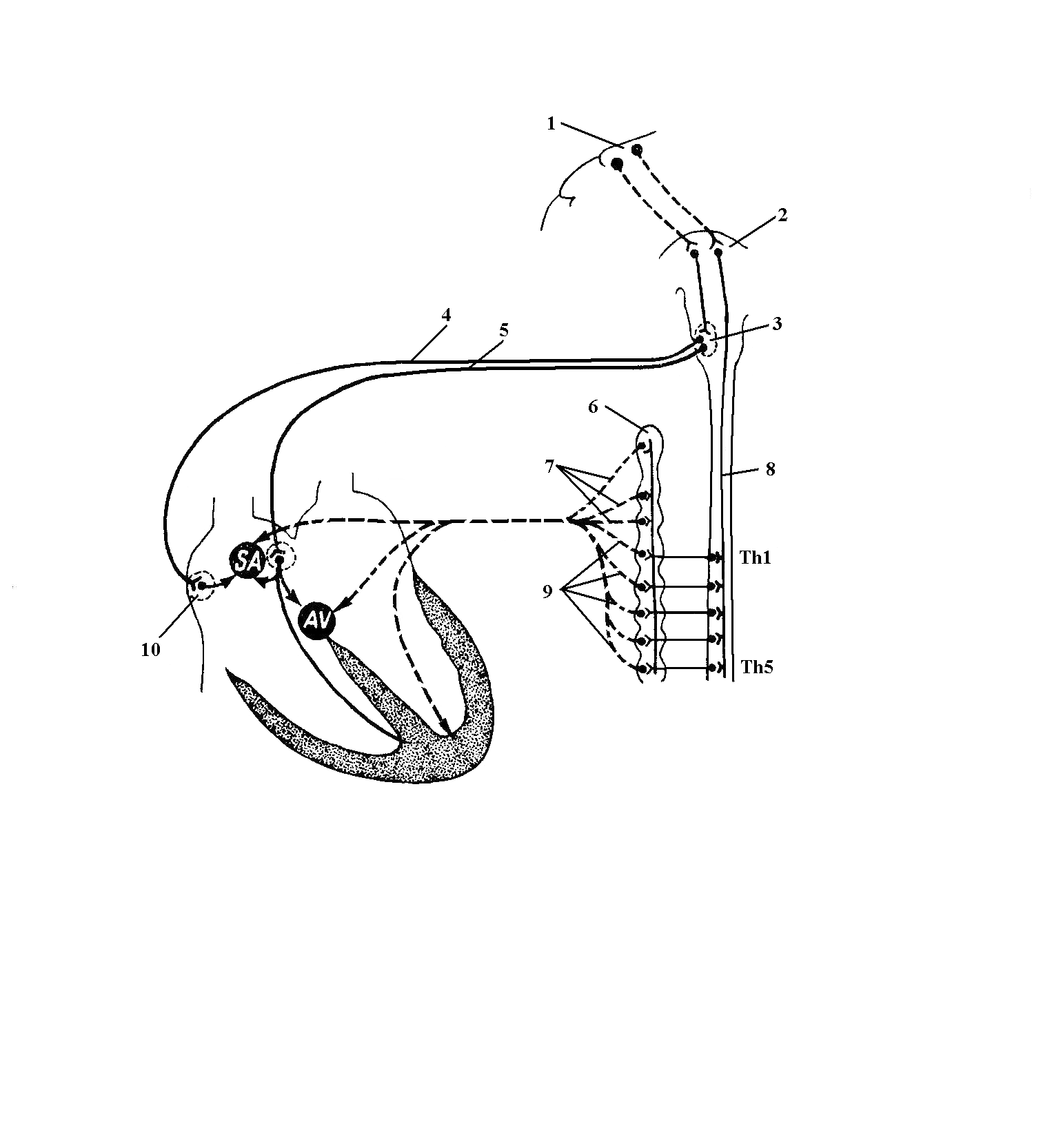 Рис. 17. Нервная регуляция деятельности
сердца:
Рис. 17. Нервная регуляция деятельности
сердца:
1 – кора головного мозга; 2 – гипоталамус; 3 – nucleus dorsalis n. vagi; 4 – n. vagus dexter; 5 – n. vagus sinister;
6 – симпатический ствол; 7 - nn. cardiaci cervicales superior, medius et inferior; 8 – задний продольный пучок;
9 - nn. cardiaci thoracici; 10 – внутриорганный парасимпатический узел;
SA – синусно-предсердный узел; AV – предсердно-желудочковый узел.
Поверхностное и глубокое сердечные сплетения по ходу правой и левой венечных артерий распространяются в подэпикардиальное сплетение, от которого отходят внутриорганные ветви, иннервирующие мышечную оболочку сердца. В сердечных сплетениях находятся разнообразные по строению и локализации группы ганглиозных клеток, морфология и физиология которых представляет большой интерес для кардиологов.
При повышении тонуса симпатических структур, принимающих участие в иннервации сердца, происходит учащение ритма сердечных сокращений (тахикардия) и усиление работы сердца. При этом артерии его, в отличие от большинства соматических и висцеральных артерий, расширяются, что соответствует возросшим энергетическим потребностям миокарда. По И.П. Павлову, здесь проявляется трофическое влияние симпатических волокон, однако существование специальных трофических нервов пока не установлено.
Парасимпатическая иннервация сердца осуществляется блуждающим нервом. Основным сегментарным центром его является дорсальное ядро продолговатого мозга. Преганглионарные волокна от нейронов этого ядра в шейном отделе блуждающего нерва отделяются в виде двух-трех верхних шейных сердечных ветвей, rr. cardiaci cervicales superiorеs, и по ходу общих сонных артерий вступают в сердечные сплетения. Нижние шейные сердечные ветви, rr. cardiaci cervicales inferiorеs, отходят от возвратного гортанного нерва (ветвь блуждающего нерва). От грудного отдела блуждающего нерва отходят грудные сердечные ветви, rr. cardiaci thoracici. Все сердечные ветви, включающие в себя преганглионарные парасимпатические волокна, заканчиваются на нервных клетках ганглиозных скоплений сердечных сплетений. Послеузловые волокна достигают мышцы и сосудов сердца.
Парасимпатическая нервная система замедляет ритм сердечной деятельности, суживает венечные артерии. Поэтому так называемая ваготония организма (повышение тонуса блуждающего нерва) сопровождается снижением частоты сердечных сокращений (брадикардия) и артериального давления, понижением обменных процессов, что характерно, например, для спящего. Исходя из этого, шейные сердечные ветви блуждающего нерва получили название n. depressor, т.е. подавляющий, угнетающий нерв.
Симпатические и парасимпатические сегментарные центры, непосредственно принимающие участие в сложной рефлекторной деятельности сердечно-сосудистой системы, находятся под постоянным влиянием и контролем со стороны вышележащих центров головного мозга. К таковым относится, прежде всего, сосудодвигательный комплекс продолговатого мозга (бульбарный вазомоторный центр). Он располагается в ретикулярной формации продолговатого мозга и физиологически подразделяется на латеральную часть, стимулирующую симпатические реакции сердца и сосудов (сердечно-ускоряющий, сосудосуживающий отдел), и медиальную часть, возбуждающую дорсальное парасимпатическое ядро блуждающего нерва. Посредством этого происходит торможение сердечной деятельности и расширение сосудов. Оба отдела вазомоторного комплекса находятся в тесном взаимодействии и координируются со стороны ретикулярной формации, гипоталамуса, коры большого мозга. Благодаря сложным морфофункциональным связям между всеми этими центрами достигается физиологическое равновесие в деятельности периферических образований симпатического и парасимпатического отделов вегетативной регуляции сердца и сосудов.
ИННЕРВАЦИЯ ОРГАНОВ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
Пищевод. Со стороны вегетативной нервной системы пищевод получает как симпатическую, так и парасимпатическую иннервацию. Следует иметь в виду, что этот орган по своему строению занимает промежуточное положение между поперечнополосатыми (произвольными) мышцами глотки и гладкой мышечной тканью стенки желудка. Поэтому он имеет соматически двигательную иннервацию (верхняя треть пищевода) и двойную вегетативную регуляцию (нижние 2/3).
Симпатические постганглионарные волокна для верхнего отдела пищевода отходят от верхнего шейного узла и в составе гортанно-глоточных ветвей вступают в пищеводное сплетение, plexus esophagealis. Симпатическая иннервация грудного отдела пищевода осуществляется за счет постганглионарных пищеводных ветвей грудных узлов (ThII-ThV) симпатического ствола. Отмечено, что большинство пищеводных нервов являются производными грудного аортального сплетения.
Спинномозговыми центрами для симпатической иннервации пищевода в целом являются участки латерального промежуточного ядра, расположенного в пределах ThI-ThIV сегментов спинного мозга. Через посредство эфферентных нейронов указанных выше симпатических узлов, они понижают или тормозят секреторную деятельность желез слизистой оболочки, суживают сосуды, расслабляют гладкую мышечную ткань, вызывая атонию и уменьшая перистальтику пищевода. При этом кардиальный вход его остается закрытым.
Парасимпатическая иннервация пищевода осуществляется блуждающим нервом, причем в этом принимают участие все его ядерные субстанции и все волокнистые компоненты. Так, мышечная оболочка верхнего отдела пищевода, как и констрикторы глотки, иннервируется за счет соматически двигательного ядра nucleus ambiguus (двойное ядро для IX и X пар черепных нервов), лежащего в продолговатом мозге. Эфферентные отростки (аксоны) клеток этого ядра в составе возвратного гортанного нерва (ветвь блуждающего нерва) непосредственно достигают поперечнополосатых мышечных слоев верхнего отдела пищевода.
Преганглионарные волокна дорсального ядра n. vagus прерываются на внутриорганных клеточных ганглиях на всем протяжении пищевода. Они иннервируют слизистую оболочку, сосуды и гладкую мускулатуру пищевода, увеличивая секрецию желез, расширяя сосуды и усиливая перистальтику. Перистальтическая волна во время произвольного акта глотания с нижнего констриктора глотки продолжается на верхний отдел пищевода («рот пищевода»), а затем рефлекторно (за счет nucleus dorsalis) переходит на гладкую мышечную ткань стенки его. При этом нижней отдел пищевода и кардиа расслабляются и пища поступает в желудок. Координированному действию различных групп мышц, участвующих в акте глотания (около 25), способствует четкая афферентная информация, поступающая в мозговые центры по чувствительным путям блуждающего, языкоглоточного и тройничного нервов.
Желудочно-кишечный тракт. Печень и поджелудочная железа. Вегетативная иннервация желудочно-кишечного тракта и пищеварительных желез обычно рассматривается как результат действия самых больших и довольно сложных внеорганных и интрамуральных нервных сплетений брюшной полости. Вполне естественно, что эти сплетения являются лишь периферическими эфферентными звеньями в цепи сложных симпатических и парасимпатических образований. Все они морфологически и функционально обусловлены вышестоящими мозговыми центрами, благодаря которым достигается согласованная регуляция двигательной и секреторной функций различных отделов пищеварительной системы.
Сегментарными центрами симпатической иннервации органов брюшной полости служат нейроны латерального промежуточного ядра спинного мозга, расположенные на протяжении от VI до XII грудного сегментов. В зависимости от иннервируемого органа, их можно подразделить на следующие группы: желудок – ThVI-ThIX, тонкая кишка – ThVII-ThX, толстая кишка – ThXI-ThXII, печень, желчный пузырь и поджелудочная железа – ThVII-ThX (рис.18).
Преганглионарные волокна от клеток латерального промежуточного ядра выходят в составе передних корешков и соответствующих указанным сегментам спинномозговых нервов и достигают грудного отдела симпатического ствола. Характерной морфологической особенностью предузловых волокон является то, что большинство из них не прерываются в грудных узлах, а проходят транзитом через симпатический ствол и отделяются от него на уровне ThVI-ThXII узлов, формируя самостоятельные большой и малый внутренностные нервы, nn. splanchnici major et minor. Последние проникают через поясничный отдел диафрагмы в брюшную полость и заканчиваются на превертебральных симпатических узлах, входящих в состав чревного, верхнего и нижнего брыжеечных сплетений.

Рис. 18. Вегетативная иннервация органов пищеварительной системы:
1 – nucleus dorsalis n. vagi; 2 – n. vagus; 3 - esophagus; 4 – gaster; 5 – intestinum tenue; 6 – intestinum crassum;
7 – парасимпатические вегетативные узлы; 8 – m. sphincter ani internus; 9 – m. sphincter ani externus;
10 – substantia intermedialateralis; 11 – nucleus motorius; 12 – ganglion coeliacum;
13 – ganglion mesentericum superius; 14 - ganglion mesentericum inferius.
Превертебральные симпатические узлы находятся у места отхождения основных ветвей брюшного отдела аорты, забрюшинно и представлены парными или непарными скоплениями периферических нейронов либо рассеянными конгломератами ганглионарных клеток. Постганглионарные волокна превертебральных узлов по ходу чревного ствола, верхней и нижней брыжеечных артерий формируют вторичные периартериальные сплетения, переходящие в органные сплетения. Так, чревное сплетение у места разветвления чревного ствола распределяется на печеночное сплетение, plexus hepaticus, иннервирующее печень, желчный пузырь, желчные пути, дуодено-пилорическую область, головку и тело поджелудочной железы, селезеночное сплетение, plexus lienalis, - для иннервации селезенки, части желудка и поджелудочной железы (хвост), желудочное сплетение, plexus gastrici.
Верхнее брыжеечное сплетение вокруг одноименной артерии и ее ветвей достигает тонкой и слепой кишки с червеобразным отростком, восходящей и поперечной ободочной кишки. Нижнее брыжеечное сплетение, на ганглионарных скоплениях которого заканчиваются преганглионарные волокна малого внутренностного нерва, иннервирует нисходящую ободочную, сигмовидную ободочную кишку и верхний сегмент прямой кишки. Следует иметь в виду, что в составе всех указанных сплетений проходят чувствительные и преганглионарные парасимпатические волокна блуждающего нерва, а также постганглионарные волокна nn. splanchnici lumbales, от поясничного отдела симпатического ствола.
По своему общему назначению симпатический отдел вегетативной нервной системы угнетает моторную деятельность желудочно-кишечного тракта и желчного пузыря, ослабляет тонус и перистальтику кишечника, вызывая атонию. При этом пилорический жом и кишечные сфинктеры сокращаются, секреторная деятельность поджелудочной железы и желез слизистой оболочки снижается, просвет кровеносных сосудов суживается.
Парасимпатическая иннервация желудочно-кишечного тракта и пищеварительных желез осуществляется за счет дорсального ядра блуждающего нерва. Преганглионарные волокна от нейронов этого ядра проделывают длинный путь в составе пищеводного сплетения, plexus esophagei, из которого далее продолжаются в виде переднего блуждающего ствола, truncus vagalis anterior, на переднюю стенку желудка и заднего блуждающего ствола, truncus vagalis posterior, на заднюю стенку желудка. При этом в составе каждого ствола находятся волокна как правого так и левого блуждающих нервов. Левый блуждающий ствол распределяется по соименным сплетениям желудка, селезенки, поджелудочной железы, желчных путей. Правый блуждающий ствол входит в состав чревного (солнечного) и брыжеечных сплетений, иннервируя тонкую кишку, поджелудочную железу, печень, толстую кишку вплоть до сигмовидной ободочной кишки.
Преганглионарные парасимпатические волокна блуждающих нервов прерываются во внутриорганных или интрамуральных узлах соответствующих органов. От узлов отходят сравнительно короткие постганглионарные волокна к гладкой мускулатуре желудочно-кишечного тракта, к секреторному аппарату пищеварительных желез, сосудам. Характерной особенностью иннервации пищеварительного тракта является наличие хорошо развитых внутриорганных нервных сплетений. Они находятся на протяжении всего желудочно-кишечного тракта – вплоть до аноректальной области.
Интрамуральные нервные сплетения представлены комплексными образованиями, состоящими из ганглионарных клеток парасимпатической природы и массы различных волокон. В кишечнике такие сплетения локализуются между продольными и циркулярными слоями мышечной оболочки (сплетение Ауэрбаха) и в подслизистой основе (сплетение Мейснера). Считается, что ганглиозные клетки межмышечного и подслизистого сплетений являются парасимпатическими образованиями, к которым подходят преганглионарные волокна конечных ветвей блуждающего нерва. Некоторые исследователи отмечают наличие здесь и нервных клеток симпатической природы. На них могут заканчиваться преганглионарные симпатические волокна, идущие в составе поясничных внутренностных нервов от симпатического ствола.
В противоположность симпатическому отделу вегетативной регуляции моторной и секреторной деятельности желудочно-кишечного тракта, парасимпатический отдел повышает тонус, частоту и амплитуду сокращений гладкомышечных образований желудка и кишечника, расслабляет сфинктеры, увеличивает секрецию всех желез, расширяет просвет кровеносных сосудов. По выражению Е.К. Сеппа, парасимпатическая система блуждающего нерва обеспечивает «стандартные функции, присущие тому или иному внутреннему органу и различным тканям». Симпатический отдел изменяет эти стандартные свойства органов, приспосабливая их деятельность к изменениям внешней и внутренней среды организма.
Из всех отделов желудочно-кишечного тракта особого внимания заслуживает иннервация прямой кишки. Как известно, в стенке ее имеется 2 сфинктера: циркулярные гладкомышечные слои заднепроходного канала (canalis analis) на протяжении 2-3 см составляют внутренний (непроизвольный) сфинктер заднего прохода, m. sphincter ani internus, а поперечнополосатые мышечные пучки тазовой диафрагмы промежности образуют наружный (произвольный) сфинктер заднего прохода, m. sphincter ani externus. Последний на 1 см черепицеобразно перекрывает внутренний сфинктер в области прямокишечно-заднепроходной линии (linea anorectalis) – места перехода слизистой оболочки кишки в кожу анального отверстия. Здесь находится большое количество разнообразных по строению и функции нервных окончаний, воспринимающих болевые, тактильные и температурные раздражения, чувство давления и растяжения.
Газы и кал, подходя к верхней трети анального канала, раздражают богато иннервируемую переходную зону слизистой оболочки, вызывают релаксацию (расслабление) внутреннего сфинктера заднего прохода, при этом наружный произвольный сфинктер сознательно сокращается или расслабляется. Регулируются эти процессы соматическим и вегетативным отделами нервной системы. В настоящее время установлено, что до 26% больных с заболеваниями прямой кишки страдают нарушениями функций анальных сфинктеров (Елисеева М.В. в соавт., 1999).
Соматически двигательная иннервация наружного сфинктера заднего прохода происходит от моторных клеток передних рогов серого вещества SII-SIV сегментов спинного мозга. Они соответствуют мозговому конусу и располагаются на уровне I-II поясничных позвонков. Отростки (аксоны) этих мотонейронов в составе передних корешков и соответствующих сакральных спинномозговых нервов достигают крестцового сплетения. Наружный произвольный сфинктер заднего прохода иннервируется половым нервом, n. pudentus, являющимся ветвью крестцового сплетения. Мотонейроны передних рогов спинного мозга находятся под постоянным двойным контролем со стороны пирамидной системы (боковой корково-спинальный тракт) и экстрапирамидной (красноядерно-спинальный тракт).
Симпатическая иннервация прямой кишки осуществляется из нижнего брыжеечного сплетения за счет периартериальных ветвей по ходу верхней прямокишечной артерии и нижнего подчревного сплетения – по ходу средней и нижней прямокишечным артериям. Спинномозговыми центрами для них являются нейроны бокового промежуточного ядра спинного мозга, находящимися в LI-LII сегментах. Симпатический отдел снижает тонус гладкомышечных слоев прямой кишки, угнетает секрецию слизистых желез и суживает кровеносные сосуды.
Парасимпатическая иннервация прямой кишки, как и всех органов полости малого таза, осуществляется сакральными парасимпатическими центрами (SII-SIV). Преганглионарные волокна от нейронов nucleus intermediolateralis проходят в составе передних корешков и 2 – 4-го крестцовых спинномозговых нервов, а затем в составе тазовых внутренностных нервов достигают интрамуральных нейронов прямой кишки. Постганглионарные волокна от них идут к гладкомышечным структурам средней оболочки и внутреннего сфинктера анального канала, усиливая тонус и перистальтику кишки и секрецию желез. Как симпатические, так и парасимпатические спинномозговые центры находятся под контролем вышестоящих вегетативных центров, благодаря чему достигается согласованная работа внутреннего и наружного сфинктеров заднего прохода.
ИННЕРВАЦИЯ ОРГАНОВ МОЧЕВОЙ СИСТЕМЫ
Почки. Сложные процессы выработки и выделения мочи, очищение крови, поддержание кислотно-щелочного равновесия и гомеостаза организма в целом находятся под постоянным контролем со стороны вегетативной нервной системы и эндокринных желез (гипоталамо-гипофизарная система, местные гормональные факторы). Причем зачастую гуморальная регуляция преобладает над нервной.
Симпатическая иннервация почек осуществляется за счет сегментарных центров, находящихся в латеральном промежуточном ядре спинного мозга на уровне 12-го грудного и 1-го поясничного сегментов. Преганглионарные волокна от нейронов этого участка ядра выходят в составе передних корешков и ThXII-LI спинномозговых нервов. Пройдя транзитом через симпатический ствол, в составе, главным образом, малых внутренностных нервов, они достигают аортопочечных симпатических узлов, ganglia aortоrenalia. Последние располагаются у места отхождения почечных артерий от брюшной части аорты. Постганглионарные волокна от аортопочечных узлов, а также отростки ганглионарных компонентов солнечного сплетения и постганглионарные волокна от поясничных узлов симпатического ствола составляют почечное сплетение, plexus renalis. По ходу почечной артерии и ее ветвей оно распределяется на пиелоуретральные, пери- и юкстаартериальные, надпочечниковые и другие ветви, иннервирующие почки, кору надпочечников, мочеточники.
Парасимпатическая иннервация этих органов осуществляется за счет ветвей заднего блуждающего ствола. Преганглионарные парасимпатические волокна, идущие от нейронов nucleus dorsalis n. vagi, заканчиваются на внутриорганных и интрамуральных ганглионарных образованиях, постганглионарные волокна которых иннервируют секреторный аппарат, сосуды и гладкомышечные структуры почек и мочевыводящих путей.
Как уже отмечалось, нервная регуляция выработки мочи, качественного и количественного состава ее, водно-солевого баланса и т.п. происходит при тесном взаимодействии с эндокринными органами. Экспериментальные исследования показывают, что стимуляция симпатического звена вегетативной нервной системы понижает фильтрацию мочи в клубочковой системе в результате сужения артериол и клубочковых капилляров под влиянием норадреналина и ренина. Преобладание симпатической иннервации приводит к снижению диуреза (греч. diuresis – образование и количество мочи), появлению в моче эритроцитов, белков и других веществ. Напротив, парасимпатический отдел стимулирует выработку мочи (полиурия), изменяет состав и солевой баланс. Более подробно механизмы нейро-гуморальной регуляции почечной деятельности рассматриваются в курсах физиологии и биохимии.
Мочевой пузырь. Мочеиспускательный канал. Процессы накопления и выведения мочи регулируются вегетативной нервной системой. Центры симпатической иннервации мочевого пузыря и рефлекса мочеиспускания находятся в латеральном промежуточном ядре спинного мозга в пределах 1 – 4-го поясничных сегментов. Преганглионарные волокна от нейронов этого ядра подходят к нижнему брыжеечному, поясничным и крестцовым узлам симпатического ствола, где передают нервный импульс на периферические эфферентные нейроны. Постганглионарные волокна от них входят в состав нижнего (тазового) подчревного сплетения, plexus hypogastricus inferior, крестцовых внутренностных нервов, nn. splanchnici sacrales, и по ходу ветвей внутренней подвздошной артерии достигают органов малого таза. В частности, по ходу верхней и нижней пузырным артериям образуется мочепузырное сплетение, plexus vesicalis.
Симпатический отдел вегетативной нервной системы расслабляет мышечную оболочку мочевого пузыря, т.е. m. detrusor vesicae, мышцу, изгоняющую мочу. При этом непроизвольный гладкомышечный сфинктер его сокращается, что дает возможность наполнения мочевого пузыря.
Парасимпатические центры для иннервации органов малого таза располагаются в боковых рогах крестцовых сегментов спинного мозга (SII-SIV). Преганглионарные волокна выходят в составе соответствующих передних корешков и крестцовых спинномозговых нервов, а затем тазовых внутренностных нервов, nn. splanchnici pelvini, присоединяются к мочепузырному нервному сплетению. Прерываются преганглионарные волокна на внутриорганных ганглионарных клетках, короткие отростки которых оплетают волокна m. detrusor и иннервируют слизистую оболочку мочевого пузыря. Они обусловливают сокращение мышечной оболочки мочевого пузыря, расслабляют сфинктер его, способствуют выведению мочи.
Рефлекс мочеиспускания обеспечивается как парасимпатическими (SII-SIV) и симпатическими спинномозговыми центрами (LI-LII), так и вышестоящими центрами коры и подкорковых образований головного мозга. Физиологическим раздражителем центров мочеиспускания является возбуждение рецепторов стенки мочевого пузыря при наполнении и расширении его. По афферентным путям эти импульсы передаются в спинномозговые вегетативные центры, а по спиноталамическим путям – в головной мозг. По нисходящим путям происходит возбуждение парасимпатических сакральных центров и торможение симпатических импульсов, в результате чего m. detrusor сокращается, непроизвольный сфинктер мочевого пузыря и произвольный сфинктер мочеиспускательного канала расслабляются и происходит волевое мочеиспускание. Таким образом, акт мочеиспускания является примером соматовегетативного действия периферических и центральных отделов соматического и вегетативного отделов нервной системы.
ИННЕРВАЦИЯ ВНУТРЕННИХ ПОЛОВЫХ ОРГАНОВ
Репродуктивная (самовоспроизведение, размножение) и эндокринная функции половых желез во всех своих постоянных или циклических проявлениях находятся под доминирующим влиянием гуморальных (гормональных) факторов. Тем не менее, общие закономерности вегетативной иннервации присущи всем половым органам, в результате чего достигается сложная нейро-гуморальная регуляция сперматогенеза и овуляции, эрекции и эякуляции и т.д. Все эти процессы регулируются и контролируются не только специальными спинномозговыми вегетативными центрами, но и, главным образом, гипоталамо-гипофизарным комплексом головного мозга.
Симпатическая иннервация внутренних половых органов осуществляется спинномозговыми центрами, заложенными в боковых рогах серого вещества спинного мозга на уровне XII грудного – I-II поясничных сегментов. Одна часть преганглионарных волокон от клеток латерального промежуточного ядра проходит транзитом через симпатический ствол и в составе малого внутренностного нерва достигает нижнего брыжеечного симпатического узла. Другая часть их спускается по межузловым ветвям симпатических стволов до поясничных и крестцовых узлов. Постганглионарные волокна от указанных узлов в составе нижнего подчревного сплетения и крестцовых внутренностных нервов (от крестцовых узлов) достигают органов малого таза, формируя вместе с преганглионарными парасимпатическими и чувствительными волокнами органные нервные сплетения.
Следует иметь в виду, что мужская и женская половые железы получают симпатическую иннервацию из аортопочечного сплетения по ходу яичковой и соответственно яичниковой артериям, вокруг которых формируются plexus testicularis et plexus ovaricus. Это связано с высоким положением половых зачатков в эмбриогенезе и последующим опусканием половых желез.
От нижнего подчревного сплетения образуются следующие вторичные сплетения:
- предстательное сплетение, plexus prostaticus, локализуется на заднебоковой поверхности железы и продолжается на мочеиспускательный канал;
- сплетение семявыносящего протока, plexus deferentialis;
- маточно-влагалищное сплетение, plexus uterovaginalis, которое концентрируется в основном в области нижнего сегмента матки и переходит на влагалище;
- пещеристое сплетение для пещеристых тел и сосудов полового члена – у мужчин и клитора – у женщин.
Центры парасимпатической иннервации половых органов, как и других органов малого таза, находятся в боковых рогах сакрального отдела спинного мозга (SII-SIV). Топографически они располагаются на уровне I-II поясничных позвонков в мозговом конусе. Преганглионарные волокна от клеток латерального промежуточного ядра проходят длинный путь в составе передних корешков конского хвоста, а затем в составе передних ветвей SII-SIV спинномозговых нервов выходят через тазовые отверстия крестца, образуя тазовые внутренностные нервы, nn. splanchnici pelvini. По ходу ветвей нижнего подчревного сплетения преганглионарные парасимпатические волокна достигают около- или внутриорганных (интрамуральных) узлов. Короткие постганглионарные волокна их иннервируют гладкомышечные оболочки, железы и сосуды половых органов.
Как отмечалось выше, нервные механизмы регуляции функций половых органов тесным образом переплетаются с гормональными факторами. Причем степень взаимодействия их зависит от функционального состояния органов. Так, установлено, что симпатический отдел в нормальных условиях (вне беременности) расслабляет мускулатуру матки, а при беременности – сокращает. Симпатические нервы способствуют процессу эякуляции (семявыбрасыванию), обеспечивая последовательность сокращения гладкомышечных образований протока придатка, семявыносящего протока, семенных пузырьков, повышению секреции предстательной и бульбоуретральных желез. Парасимпатическая иннервация обеспечивает эрекцию полового члена и клитора, которая осуществляется за счет парасимпатических волокон, проходящих к пещеристым телам в составе внутреннего полового нерва (эректор Экарта).
Обобщенные данные локализации центральных и периферических нейронов эфферентного звена рефлекторной дуги вегетативной нервной системы представлены в таблице 2.
|
Таблица 2 |
||||
|
Локализация центральных и периферических нейронов эфферентного звена вегетативной нервной системы |
||||
|
Иннервируемый орган |
Симпатический отдел |
Парасимпатический отдел |
||
|
Сегментарный центр |
Вегетативный узел |
Сегментарный центр |
Вегетативный узел |
|
|
Мышца, расширяющая зрачок |
Латеральное промежуточное ядро CVIII, ThI-ThII сегментов спинного мозга |
Верхний шейный узел симпатического ствола |
- |
- |
|
Мышца, суживающая зрачок и ресничная мышца |
- |
- |
Добавочное ядро глазодвигатель-ного нерва (средний мозг) |
Ресничный узел |
|
Слезная железа |
Латеральное промежуточное ядро ThI-ThII сегментов спинного мозга |
Верхний шейный узел симпатического ствола |
Верхнее слюноотделитель-ное ядро (мост) |
Крылонебный узел |
|
Околоушная слюнная железа |
Латеральное промежуточное ядро ThI-ThII сегментов спинного мозга |
Верхний шейный узел симпатического ствола |
Нижнее слюноотделитель-ное ядро (продолговатый мозг) |
Ушной узел |
|
Поднижнече-люстная и подъязычная слюнные железы
|
Латеральное промежуточное ядро ThI-ThII сегментов спинного мозга |
Верхний шейный узел симпатического ствола |
Верхнее слюноотделитель-ное ядро (мост) |
Крылонебный узел |
|
Таблица 2 (продолжение) |
||||
|
Иннервируемый орган |
Симпатический отдел |
Парасимпатический отдел |
||
|
Сегментарный центр |
Вегетативный узел |
Сегментарный центр |
Вегетативный узел |
|
|
Глотка, гортань |
Латеральное промежуточное ядро ThI-ThII сегментов спинного мозга |
Верхний шейный узел симпатического ствола |
Дорсальное ядро блуждающего нерва (продолговатый мозг) |
Интрамураль-ные узлы |
|
Трахея, бронхи, легкие |
Латеральное промежуточное ядро ThII-ThVI сегментов спинного мозга |
Звездчатый и верхние грудные узлы симпатического ствола |
Дорсальное ядро блуждающего нерва (продолговатый мозг) |
Интрамураль-ные узлы |
|
Сердце |
Латеральное промежуточное ядро ThI-ThV сегментов спинного мозга |
Шейные и верхние грудные узлы симпатического ствола |
Дорсальное ядро блуждающего нерва (продолговатый мозг) |
Около- и внутриорган-ные узлы |
|
Пищевод |
Латеральное промежуточное ядро ThI-ThIV сегментов спинного мозга |
Верхние грудные узлы симпатического ствола |
Дорсальное ядро блуждающего нерва (продолговатый мозг) |
Интрамураль-ные узлы |
|
Желудок, тонкая кишка, толстая кишка, печень, поджелудочная железа |
Латеральное промежуточное ядро ThVI-ThXII сегментов спинного мозга |
Превертебраль-ные узлы: чревный, верхний и нижний брыжеечные |
Дорсальное ядро блуждающего нерва (продолговатый мозг) |
Интрамураль-ные узлы Внутриорган-ные узлы |
|
Прямая кишка |
Латеральное промежуточное ядро ThXII– LIII сегментов спинного мозга |
Нижний брыжеечный узел |
Боковые рога серого вещества спинного мозга на уровне SII-SIV сегментов |
Интрамураль-ные узлы |
|
Почки |
Латеральное промежуточное ядро ThXII- LI сегментов спинного мозга |
Аортопочечные узлы |
Дорсальное ядро блуждающего нерва (продолговатый мозг) |
Интрамураль-ные узлы Внутриорган-ные узлы |
|
Мочевой пузырь и мочеиспуска-тельный канал |
Латеральное промежуточное ядро LI-LIV сегментов спинного мозга |
Нижний брыжеечный, поясничные и крестцовые узлы |
Боковые рога серого вещества спинного мозга на уровне SII-SIV сегментов |
Интрамураль-ные узлы |
|
Половые органы |
Латеральное промежуточное ядро ThXII- LII сегментов спинного мозга |
Нижний брыжеечный, поясничные и крестцовые узлы |
Боковые рога серого вещества спинного мозга на уровне SII-SIV сегментов |
Около- и внутриорган-ные узлы |
АФФЕРЕНТНОЕ ЗВЕНО ВЕГЕТАТИВНОЙ РЕФЛЕКТОРОЙ ДУГИ И ЧУВСТВИТЕЛЬНАЯ ИННЕРВАЦИЯ ВНУТРЕННИХ ОРГАНОВ
Вопрос о чувствительной иннервации внутренних органов в настоящее время не вызывает сомнений. Во всех полых и паренхиматозных органах, сосудах, кроме эффекторных нейро-тканевых окончаний, имеется большое количество специализированных рецепторов (интерорецепторов), которые по своей морфологии и физиологии подразделяются на несколько групп:
- барорецепторы воспринимают давление биологических жидкостей (например, давление крови в сосудах);
- механорецепторы регистрируют степень растяжения органа;
- хеморецепторы чувствительны к изменениям состава и концентрации определенных веществ в жидкостных средах организма;
- осморецепторы активируются при изменении осмотического давления крови, тканевой жидкости межклеточных и интерстициальных пространств.
Информация от интерорецепторов передается по афферентным путям в центральную нервную систему, в том числе и в кору большого мозга, однако многие раздражения не осознаются, сознательно не воспринимаются. Например, снижение парциального давления кислорода не ощущается и проявляется лишь через функциональные изменения дыхательной и сердечно-сосудистой систем. Активация механорецепторов желудка вызывает чувства наполнения и сытости, а раздражение осморецепторов – чувство жажды. Раздражение интерорецепторов почек и мочеточников обусловливает резкие боли. Растяжение стенки мочевого пузыря является предвестником акта мочеиспускания и т.д.
Среди проблем интероцептивной чувствительности наиболее важными, но недостаточно выясненными остаются вопросы висцеральной болевой чувствительности. Прежние представления об отсутствии болевой чувствительности внутренних органов не согласуются с повседневной общеврачебной практикой: при большинстве заболеваний внутренних органов возникают наиболее упорные и мучительные боли (сердца, желудка, кишечника и т.д.). В настоящее время механизмы происхождения висцеральных болей отчасти раскрыты и продолжают интенсивно изучаться.
В частности, установлены 2 типа болевых рецепторов (ноцицепторов). Это свободные нервные окончания миелиновых А-дельта волокон, проводящие нервный импульс со скоростью от 6 до 30 м/сек. Такие рецепторы находятся в коже, слизистой оболочке полости носа и рта, возбуждаются сверхинтенсивными раздражителями (укол, ожог) и быстро проводят раздражение.
Другой тип ноцицепторов представлен некапсулированными клубочковыми окончаниями немиелиновых С-волокон, скорость проведения импульса по ним мала – 0,5–2м/сек. Такие ноцицепторы характерны для внутренних органов. Они отличаются полимодальностью, т.е. реагируют как на механические, так и температурные и химические раздражения и активируются продуктами распада тканей при различных органических поражениях органов («тканеповреждающие рецепторы»).
Проводящие пути всех видов интерорецепции обычно отождествляются с афферентными путями соматических рефлекторных дуг.
Первые нейроны всех видов афферентации внутренних органов находятся на периферии в чувствительных узлах спинномозговых и черепных нервов (V, VII, IX, X пары). Периферические отростки псевдоуниполярных клеток спинномозговых узлов, пройдя в составе белых соединительных ветвей, затем в составе шейных и грудных внутренностных нервов (для органов грудной полости), большого и малого внутренностных нервов (для органов брюшной полости) и тазовых внутренностных нервов (для органов малого таза), связываются с механо-, баро- и хеморецепторами внутренних органов, а также с ноцицепторами (болевыми рецепторами).
Центральные отростки клеток спинномозговых узлов в составе задних корешков входят в спинной мозг и разделяются на восходящие и нисходящие пучки, охватывая несколько сегментов. Кроме того, по современным данным (Кривошапкин А.Л., 2005), до 30% центральных отростков спинальных ганглиев может проходить через передние корешки. Этим объясняется безуспешность снятия болей путем выборочного пересечения тонких безмиелиновых ноцицептивных волокон задних корешков спинного мозга.
В спинном мозге центральные волокна, несущие интероцептивную чувствительность, заканчиваются на вторых нейронах, лежащих в промежуточной зоне серого вещества, кзади от латерального промежуточного ядра. Кроме того, часть волокон подходит к этому ядру, замыкая короткую симпатическую рефлекторную дугу в пределах спинного мозга, которая осуществляет автоматическое управление деятельностью внутренних органов.
Отростки вторых нейронов интероцептивной чувствительности малочисленные, поэтому не образуют специальных трактов и пучков. Они присоединяются к другим восходящим путям заднего и бокового канатиков спинного мозга и достигают заднелатеральных и заднемедиальных ядер зрительного бугра. От таламуса по отросткам третьих нейронов импульсы в составе таламокортикальных путей через середину задней ножки внутренней капсулы поступают в нижний отдел постцентральной извилины, а также в премоторную зону лобной доли и височную долю большого мозга.
Стволовая афферентная иннервация (по Гайворонскому И.В., 2000) внутренних органов происходит за счет чувствительных волокон лицевого, языкоглоточного и блуждающего нервов. Как и в спинномозговом отделе, периферические отростки псевдоуниполярных клеток, лежащих в соответствующих узлах этих нервов (узел коленца, верхний и нижний узлы IX и X пар), связаны с интерорецепторами органов, а центральные отростки их заканчиваются на клетках ядра одиночного пучка, общего для VII, IX и X пар черепных нервов. Отростки вторых нейронов в составе ядерно-таламического тракта достигают зрительного бугра и переключаются на третьи нейроны базальных ядер таламуса. Отростки третьих нейронов по классическому таламо-кортикальному пути, через среднюю часть задней ножки внутренней капсулы, подходят к корковому ядру висцерального анализатора – нижнему отделу постцентральной извилины.
В коре происходит анализ полученной информации, интеграция вегетативных и соматических рефлексов и передача соответствующих сигналов на подкорковые центры вегетативной регуляции. По ассоциативным путям импульсы от гипоталамо-гипофизарного комплекса достигают непосредственно головных и спинномозговых центров симпатического и парасимпатического отделов.
В нормальных условиях деятельности внутренних органов интероцептивная чувствительность в большинстве случаев не формирует в коре каких-либо конкретных, осознанных ощущений, т.е. находится вне порога нашего сознания. Это объясняется слишком слабым, подпороговым, действием раздражителей внутренней среды на интерорецепторы и ограниченной проекцией висцеральной чувствительности в коре головного мозга.
Афферентные интероцептивные раздражения приобретают особую роль в патологических условиях, когда они становятся сверхпороговыми. В таких случаях на первый план выступает болевой синдром как защитная реакция организма, связанная с запредельным раздражением и повреждением тканей. Висцеральная боль возникает вследствие быстрого сокращения или спазма мышечной оболочки органа, чрезмерного растяжения стенки, сужения кровеносных сосудов и т.п. Отключение патологических импульсов на различных уровнях висцерального анализатора с помощью фармакологических средств лежит в основе терапии тех или иных заболеваний внутренних органов.
ЗАКЛЮЧЕНИЕ
Всестороннее и более глубокое рассмотрение вопросов анатомии и физиологии вегетативной нервной системы убеждает нас в том, что деление единой нервной системы на соматический и вегетативный отделы является весьма условным. Оно может означать лишь разделение точек приложения периферических иннервационных механизмов при сохранении и преобладании интегрирующей роли высших центров.
Как известно, в основе возникновения и специализации нервных структур, как и других клеточных образований, лежит универсальное свойство живой ткани – раздражимость, т.е. закономерная реакция тканей на различные факторы внешней и внутренней среды. В результате этого наружные покровы тела становятся мощным «биологическим экраном», воспринимающим и передающим в центральную нервную систему многочисленные раздражения внешней среды (механические, физические, температурные и др.), формируются и специальные органы чувств (зрения, слуха, обоняния, вкуса). Вся эта соматическая афферентация передается не только на специальные корковые и подкорковые центры соматической рефлекторной дуги, но и по типу сомато-висцеральных рефлексов замыкается на надсегментарных вегетативных центрах, имеющих отношение к иннервации внутренних органов. Поэтому в ответ на внешние раздражения меняется деятельность сердечно-сосудистой и дыхательной систем, пищеварительных и мочевых органов, желез внешней и внутренней секреции. Все это обусловливает адаптацию организма к меняющимся условиям внешней среды, ведет к изменению трофических процессов в организме.
Аналогичные механизмы лежат в основе происхождения и развития богатых иннервационных структур внутренних органов. Причем количество источников, распределение и структура их находятся в прямой зависимости от строения и функции органов. Интероцептивный рецепторный аппарат воспринимает и передает в центральную нервную систему информацию о всех изменениях внутренней среды организма (механические, химические, барометрические и другие раздражения). При этом информация о состоянии внутренних органов и трофических процессов, опять-таки, поступает не только в центры вегетативной регуляции, но и обменивается между корковыми и подкорковыми центрами соматического отдела (висцеро-соматические рефлексы).
Таким образом, в основе строения соматического и вегетативного отделов нервной системы лежат единые принципы возникновения и функциональной деятельности обеих систем. Морфологические различия между ними состоят лишь в том, что вегетативный отдел характеризуется повсеместностью распространения во всех органах и тканях. Однако следует иметь в виду, что распределение симпатических и парасимпатических волокон в них может быть различным.
В отличие от симпатических нервных структур, которые диффузно распределены по всему телу, парасимпатические контролируют более ограниченные соматические и висцеральные территории. Например, краниальный отдел вегетативной нервной системы обеспечивает парасимпатическую иннервацию органов головы, грудной и брюшной полостей, сакральный отдел – органов малого таза. Периферические артерии тела и потовые железы лишены парасимпатической иннервации. Вариации резистентности (тонуса) сосудов находятся под исключительным контролем симпатических нервов. В селезенке и почке также отмечается преобладание симпатической иннервации, тогда как пищеварительный тракт находится под преобладающим влиянием парасимпатического компонента (n. vagus) вегетативной нервной систем.
Другой важной особенностью строения соматического и вегетативного отделов нервной системы являются различия в локализации эфферентного звена рефлекторной дуги. Если в соматическом отделе эфферентные нейроны (мотонейроны) находятся в стволовой части головного мозга и в передних рогах спинного мозга, то эфферентное звено вегетативной нервной системы имеет двухчленный характер, причем последний нейрон его находится на периферии - в вегетативных узлах или ганглиозно-сетчатых образованиях вне- и внутриорганных сплетений. В связи с меньшей миелинизацией вегетативных волокон и сравнительно небольшой скоростью проведения нервного импульса реакция внутренних органов возникает медленно, но может быть длительной.
Вегетативная нервная система в целом и за счет эфферентного симпатического и парасимпатического отделов ее обеспечивает нормальную жизнедеятельность всех висцеральных органов, а также адаптационно-трофическую функцию всего тела. Поэтому поражение вегетативных нейронов или отростков их (проводящих путей) ведет к нарушению функций внутренних органов (моторных, секреторных), с появлением так называемых висцеро-дисфункциональных синдромов. Дальнейшие нарушения трофической функции вегетативной нервной системы приводят к возникновению висцеро-дистрофических синдромов, к органическим изменениям внутренних органов, т.е. соматическим заболеваниям.
Выявление патофизиологических механизмов заболеваний человека требует глубоких знаний не только развития, строения и функции каждого органа, но и его регулирующих систем. Все это имеет прямое отношение к проблемам медицинской практики, к задачам наиболее раннего выявления нарушений гомеостаза и дисфункций внутренних органов. Яркими примерами тому могут служить дисфункции (висцеропатии) различных внутренних органов вертеброгенной этиологии: тесная топическая связь корешковых структур вегетативных проводников, узлов симпатического ствола с позвоночным столбом обусловливает поражение их при дегенеративно-дистрофических изменениях межпозвоночных дисков (остеохондроз).
Так, при остеохондрозе шейного отдела позвоночного столба часто возникают сердечные расстройства с нарушением ритма, появлением длительных или приступообразных болей в области сердца, за грудиной.
При патологии грудного отдела позвоночного столба нарушается дыхательная функция с болезненностью и уменьшением подвижности ребер, что может привести к застойным явлениям в легких и развитию пневмонии. При поражении шейно-грудных сегментов спинного мозга наблюдаются спазмы пищевода, нарушается пищеводный акт глотания пищи, появляются сдавливающие боли за грудиной. Поражение средних и нижних грудных корешков и симпатических узлов сопровождается секреторными и двигательными нарушениями со стороны различных отделов желудочно-кишечного тракта, запорами или, наоборот, жидким стулом, тошнотой, с последующим появлением органических дистрофических нарушений (язва желудка, двенадцатиперстной кишки).
Компрессия поясничных сегментов спинного мозга и корешков конского хвоста ведут к расстройствам мочевых и половых органов: появлению болей в виде типичной почечной колики, нарушению функции мочевого пузыря и акта мочеиспускания (задержка или недержание мочи), снижению мужской потенции, нарушению менструального цикла у женщин.
Диагностика вертеброгенных нарушений функций внутренних органов, особенно кардиальных, значительно затруднена. Она усложняется еще и тем, что внутренние органы не имеют четкой сегментарной иннервации, характерной для соматического отдела нервной системы. Поэтому все функциональные и патологические реакции не ограничиваются одним органом или областью. Они могут иррадиировать на другие, казалось бы, функционально малосвязанные органы. Этому способствует принцип мультипликации преганглионарных вегетативных волокон, которые многократно ветвятся и заканчиваются не десятках нейронов вегетативных узлов, являющихся источником иннервации многих органов и сосудов.
Приведенные в учебном пособии теоретические и некоторые прикладные аспекты морфологии и физиологии вегетативной нервной системы свидетельствуют о том, что при выяснении патогенетических механизмов тех и или иных заболеваний необходим более тщательный анализ состояния иннервационных структур каждого органа и трофических процессов в нем. Вполне естественно, что для дальнейшего развития учения о вегетативной нервной системе морфологические методы имеют ограниченные возможности. Для этого необходимы совместные усилия морфологов, физиологов, биохимиков, фармакологов, патологов и клиницистов.