

Гликолиз и глюконеогенез – взаимосвязанные процессы.
Глюконеогенез и гликолиз протекают в основном в цитоплазме. Так как в процессе глюконеогенеза глюкоза синтезируется, а в гликолизе расщепляется, очевидно, что оба процесса должны контролироваться взаимосвязанно. В противном случае их работа была бы бесполезной.
Гормональная регуляция. Повышение уровня глюкозы в крови приводит к выбросу инсулина α-клетками поджелудочной железы. Инсулин стимулирует поступление глюкозы в ткани и утилизацию её (в реакциях гликолиза и других) клетками организма, следствием чего является снижение концентрации глюкозы в крови. Гипогликемия провоцирует поступление в кровоток глюкагона (гормон β-клеток поджелудочной железы) и глюкокортикоидов, которые являются мощными активаторами глюконеогенеза. Следствием стимулированного глюконеогенеза становится повышение концентрации глюкозы в крови (рис 6.11)
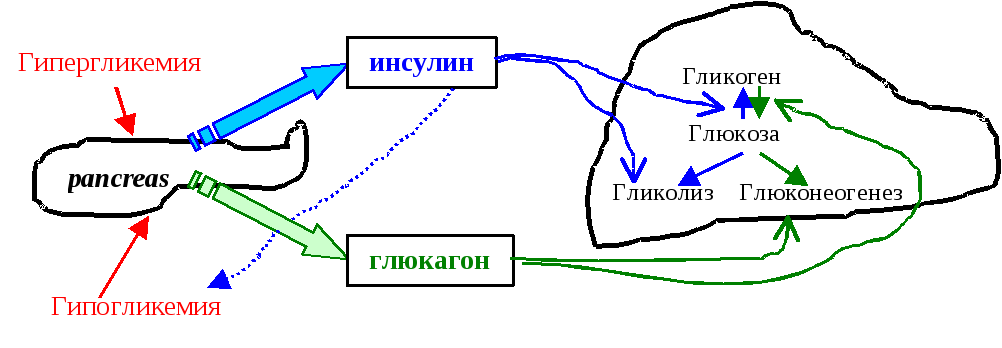
Рис. . Регуляция гликолиза и глюконеогенеза гормонами поджелудочной железы.
Внутриклеточная регуляция. Регуляция глюконеогенеза тесно связана с регуляцией гликолиза. В условиях энергетической недостаточности активируются скорость контролируемые реакции гликолиза. Напротив, глюконеогенез требует затрат энергии и протекает в условиях, при которых поддерживается достаточный уровень АТФ в клетке.
Регуляция осуществляется путём аллостерического контроля ферментов, которые для этих двух путей различны.
-
Активности гексокиназы и глюкозо-6-фосфатазы регулируются уровнем глюкозо-6-фосфата: гексокиназа им ингибируется, а фермент глюконеогенеза (т.е.глюкозо-6-фосфатаза) активируется.
-
Для пируваткиназы и пируваткарбоксилазы аллостерческим эффектором является ацетил-КоА, однако если для фермента глюконеогенеза он является положительным модулятором, то активность пируваткиназы ацетил-КоА, напротив, ингибирует.
-
Главным аллостерическим регулятором двух взаимосвязанных путей – гликолиза и глюконеогенеза – является фруктозо-2,6-дифосфат: увеличение его концентрации активирует ключевой фермент гликолиза – фосфофруктокиназу-1; снижение его концентрации активирует конкурирующую реакцию – образование фруктозо-6-фосфата, т.е. приводит к усилению глюконеогенеза (рис.).


Рис. 5.Аллостерический контроль фосфофруктокиназной активности.
Фруктозо- 2,6-бифосфат синтезируется бифункциональным ферментом - фосфофруктокиназой-2 (ФФК-2). Фермент состоит из 2-х идентичных субъединиц, каждая из которых имеет собственный каталитический центр. ФФК-2 активируется цАМФ, увеличение концентрации которого приводит к фосфорилированию фермента (точнее, ФФК-2 фосфорилируется цАМФ-зависимой протеинкиназой А). Фосфорилированный фермент проявляет фосфатазную активность – образуется фруктозо-2,6-дифосфат. Уменьшение концентрации цАМФ вызывает дефосфорилирование ФФК-2 – растёт его киназная активность, вследствие чего образование фруктозо-2,6-бифосфата снижается.
Подчеркнём, что повышение концентрации цАМФ в печени обуславливается действием глюкагона и адреналина; напротив, под действием инсулина она снижается. Следовательно, активируемое адреналином и глюкагоном повышение фосфорилирования ФФК-2 уменьшит образование фруктозо-2,6-бифосфата, дефицит которого приведёт к угнетению активности ФФК-1 и ингибированию гликолиза. Инсулин, активируя синтез фруктозо-2,6-дифосфата (за счёт падения концентрации цАМФ), тем самым будет активировать ФФК-1 и стимулировать гликолиз.
Фруктозо-2,6-бифосфатаза жёстко ингибируется фруктозо-6-фосфатом.
Глюкагон репрессирует транскрипцию пируваткиназы и активирует транскрипцию фосфоэнолПВКкарбоксикиназы. Инсулин репрессирует транскрипцию фосфоэнолПВКкарбоксикиназы.