

Макроэргические соединения
В живом организме нет прямого перехода энергии от источника (энергогенерирующего процесса) к потребителю (биологическому преобразователю энергии). Непосредственное использование потенциальной химической энергии, содержащейся в молекулах пищевых веществ, невозможно потому, что при разрыве внутримолекулярных связей выделяется такое огромное количество энергии, что она не может быть аккумулирована внутриклеточными ферментными системами и способна вызвать повреждение этих систем. Для процессов, совершающихся в живом организме, характерно ступенчатое освобождение свободной энергии с одновременным преобразованием ее в химическую энергию макроэргических соединений или электрохимический потенциал на клеточных мембранах.
Макроэргические соединения это соединения, содержащие макроэргическую связь. Макроэргическими называются химические связи, свободная энергия гидролиза которых составляет не менее 21 кДж/моль. Эти связи обозначают знаком ~ (тильда). Говоря об энергии макроэргических связей, в биохимии имеют в виду не действительную энергию ковалентной связи между атомами, как это принято в физической химии, а лишь разность между значениями свободной энергии (∆G) исходных реагентов и продуктов реакций гидролиза макроэргических соединений. «Энергия связи» в этом смысле, строго говоря, не локализована в данной связи, а характеризует реакцию в целом. Поэтому правильнее говорить не о возникновении макроэргической связи, а об образовании в процессе химического сопряжения нового макроэргического соединения.
Универсальной формой запасания свободной энергии для всего живого мира является энергия макроэргических связей АТФ: все преобразования энергии в процессах жизнедеятельности осуществляются через аккумуляцию энергии в этих связях и её использование при их разрыве. Значение ∆G для этих реакций представляет собой как бы «биологический квант» энергии, т.к. все преобразования энергии в организмах происходят порциями, примерно равными ∆G. При ферментативном гидролизе АТФ в клетке отщепляющаяся фосфатная группа переносится на субстрат, запас энергии в котором оказывается в результате больше, чем в исходном соединении.
Величина изменения свободной энергии в реакциях переноса зависит как от природы переносимой группы, так и от природы молекулы акцептора. При сравнении потенциала переноса групп различных соединений необходимо пользоваться единым стандартным акцептором. В качестве такого акцептора обычно принимают молекулу воды и выражают потенциал переноса в виде свободной энергии реакции гидролиза данного соединения.
Потенциал переноса измеряется в кДж на 1 моль групп донора, переносимых на стандартный акцептор при стандартных (одномолярных) концентрациях. В таблице 1 представлены величины ∆G0 (при рН=7, Т=250С) основных макроэргических соединений и органических соединений с нормальными связями.
Таблица 1. Свободная энергия гидролиза некоторых соединений.
|
Соединение |
∆G0 , кДж/моль |
|
Фосфоенолпируват |
-61,9 |
|
1,3-дифосфоглицерат |
-49,4 |
|
Карбамоилфосфат |
-51,5 |
|
Ацетилфосфат |
-43,15 |
|
Креатинфосфат |
-37,7 |
|
Сукцинил-КоА |
-33,5 |
|
Ацетил-КоА |
-31,4 |
|
АТФ (до АМФ) |
-31,8 |
|
АТФ (до АДФ) |
-31,0 |
|
АДФ (до АМФ) |
-28,3 |
|
Глюкозо-1-фосфат |
-20,7 |
|
Фруктозо-6-фосфат |
-15,8 |
|
Глюкозо-6-фосфат |
-13,8 |
|
Глицерофосфат |
10,0 |
Сопоставляя величины потенциалов переноса, можно выяснить, является ли определенная комбинация реакций термодинамически вероятной. В условиях стандартных концентраций происходит самопроизвольный перенос групп от соединений, у которых потенциал переноса высок, к соединениям, у которых он ниже. Так, АТФ, находящаяся в середине шкал значений потенциала переноса групп, способна акцептировать фосфорную группу от тех соединений, которые в реакциях энергетического метаболизма выполняют роль первичных макроэргов и имеют более высокий потенциал переноса. Вместе с тем АТФ может переносить фосфорные группы на молекулы соединений, относящихся к «низкоэнергетическим» фосфатам, у которых потенциал переноса групп ниже, чем у АТФ.
Большинство макроэргических соединений представляют собой ангидриды фосфорной кислоты и какой-либо другой кислоты, или фосфорные эфиры енолов. Первично возникающие в ходе реакций химического сопряжения макроэргические соединения отличаются высокой нестабильностью в водной среде. Они легко подвержены гидролизу, что ведет к рассеиванию энергии окисления в виде тепла. Поэтому для того, чтобы сохранить энергию и использовать ее в процессах, идущих с потреблением энергии, первично возникшая макроэргическая связь должна быть перенесена на молекулу соединения, обладающего достаточно высокой устойчивостью в водной среде. В большинстве случаев макроэргические фосфорные соединения при участии фосфотрансфераз переносят фосфатную группу на молекулу АДФ, выполняющую в клетке роль универсального акцептора высокоэнергетического фосфата. Образующаяся в итоге подобных реакций переноса АТФ выступает затем как специфический донор фосфатных групп в других ферментативных реакциях.
АТФ
Молекулярное строение и химические свойства АТФ соответствуют ее функции промежуточного аккумулятора и переносчика энергии. Молекула АТФ состоит из органического основания аденина, углевода рибозы и трех остатков фосфорной кислоты, из которых пирофосфатные группы являются макроэргическими. Формулу АТФ можно записать так: аденин - рибоза - фосфат ~ фосфат ~ фосфат.
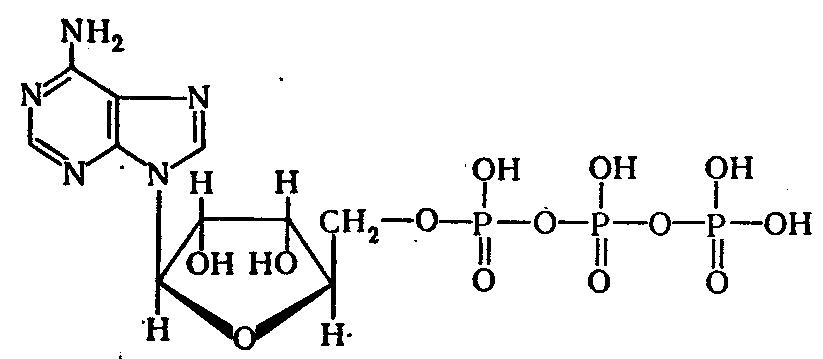
Соединение аденина и рибозы - это аденозин, а соединение аденозина с первым остатком фосфорной кислоты - адениловая или аденозинмонофосфорная кислота (АМФ). Адениловая кислота относится к группе нуклеотидов, входящих в состав нуклеиновых кислот. В молекуле АТФ по главной цепи -Р-О-Р-О- происходит накопление положительных зарядов, электростатическое отталкивание которых вносит определенный вклад в образование макроэргических связей. Наличие нескольких отрицательных зарядов вокруг центральной цепи положительных зарядов играет защитную роль, благодаря чему молекула АТФ способна накапливать энергию при одновременной кинетической устойчивости в водной среде. В то же время молекулы воды легко атакуют пирофосфатную структуру, поскольку электростатическое отталкивание отрицательно заряженных фосфатных групп способствует их разобщению. Это сильно выраженное стремление к отщеплению концевых фосфатных групп АТФ представляет собой движущую силу в ферментативных реакциях переноса, которые сопровождаются фосфорилированием других соединений за счет АТФ.
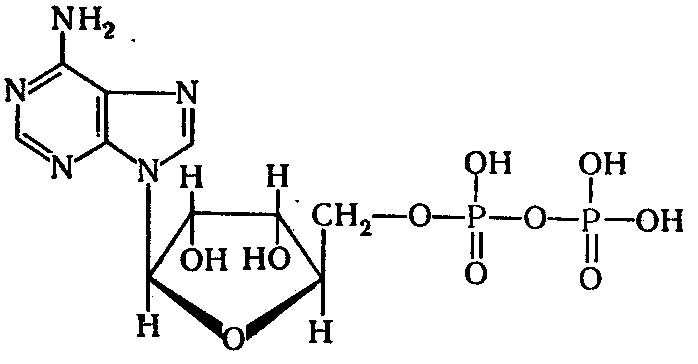
Связь между кислородом и фосфором в адениловой кислоте - обычная сложноэфирная связь, при гидролитическом отщеплении которой освобождается энергия около 13 кДж/моль. При гидролизе АТФ с образованием молекулы ортофосфорной кислоты и АДФ в стандартных условиях освобождается энергия около 31 кДж/моль:
АТФ + Н2О = АДФ + Н3РО4
+ Н2О =
АТФ
ОН
│
+ НО ─ Р ─ ОН
║
О
АДФ
Количество освобождаемой энергии зависит от рН среды, концентрации неорганического фосфата, а также соотношения концентраций АТФ и АДФ в клетке. При отщеплении второй фосфатной группы образуется аденозинмонофосфат и выделяется примерно такое же количество энергии:
АДФ + Н2О = АМФ + Н3РО4
Возможен также и другой путь расщепления, при котором от молекулы АТФ отщепляются сразу две фосфатные группы (пирофосфат) и образуется АМФ, ∆G0 = - 31,8 кДж/моль:
АТФ + Н2О = АМФ + Н4Р2О7
Молекула АМФ может снова фосфорилируоваться до АДФ второй молекулой АТФ при участии особого фермента аденилаткиназы, и образующийся пирофосфат гидролизуется пирофосфатазой до ортофосфорной кислоты:
АМФ+АТФ = 2АДФ + Н4Р2О7
Наличие двух путей гидролитического расщепления АТФ обусловливает дополнительную гибкость адениловой системы в функции посредника между процессами аккумуляции и использования энергии.
АТФ может функционировать в клетках не только как фосфорилирующий агент, переносящий фосфат на соответствующий акцептор, но и как донор, переносящий на акцептор неорганический пирофосфат, и как аденилирующий агент, переносящий на акцептор свою аденилатную группу (например, при образовании аминоациладенилатов).
При фосфорилировании АДФ в процессах, сопряженных с биологическим окислением, должна быть затрачена энергия, достаточная для создания необходимого потенциала переноса групп. Величину энергии зависит не только от температуры давления, но и от рН и от концентрации реагентов, которая может быть различной в разных отделах клетки. Так, для концентрации в цитоплазме величина свободной энергии гидролиза АТФ составляет 40 кДж/моль, а в митохондриях, где в основном образуется АТФ, - 60 кДж/моль.
Аденилатная система клетки включает: АДФ, АТФ и АМФ. Сумму этих компонентов в клетке практически постоянная величина. Энергетический статус клетки в каждый определенный момент зависит от относительных концентраций всех трех компонентов. Если вся система представлена в форме АТФ, то клетка находится в максимально высоком энергетическом состоянии; если же в форме АМФ, то клетка лишена богатых энергией компонентов и находится в низшем энергетическом состоянии. Для оценки энергетических состояний пользуются показателем энергетического заряда аденилатной системы, который равен половине общего числа макроэргических фосфатных групп, приходящихся на остаток аденозина.
Максимальный энергетический заряд, когда вся аденилатная система представлена АТФ, равен единице, а минимальный, когда единственным компонентом ее является АМФ, - нулю.
В живой клетке величина «энергетического заряда» поддерживается пределах от 0,7 до 0,9. Изменение энергетического заряда приводит к большим изменениям скорости процессов, замедлению или ускорению действия ферментов. Таким образом, система адениловых нуклеотидов осуществляет регуляцию процессов, ответственных за поставку и утилизацию энергии в клетке, и поддерживает оптимальный уровень функционирования, требующий наименьших затрат энергии.
В клетках есть несколько метаболических путей, ведущих к синтезу АТФ в сопряженных реакциях. Эти пути различаются по характеру катаболических превращений, приводящих к образованию доноров электронов, по типу окислительного процесса, в ходе которого высвобождается потенциальная свободная энергия, и по способу сопряжения энергоподающей реакции с образованием макроэргических соединений. Каждый из этих процессов приспособлен к обработке определенного вида «топлива», и каждый из них имеет свои пределы эффективности преобразования энергии окисления при синтезе АТФ.