

-
Єдиний нейрогуморальний механізм регуляції
???
Білет 13
-
Цтк і механізми, що контролюють його у мозку
???
-
Катаболізм піримідинових нуклеотидів
Р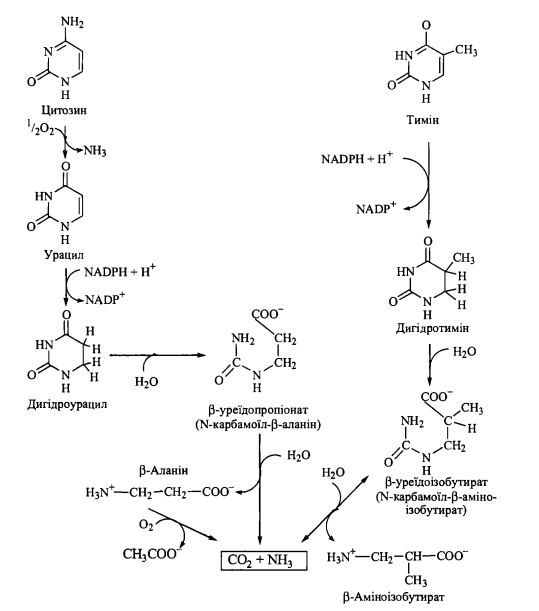 озщеплення
піримідинових нуклеотидів Початкові
етапи катаболізму піримідинових
нуклеотидів, як і в разі пуринових
нуклеотидів, полягають у відщепленні
рибозофосфату з подальшим окисленням
утворених піримідинів. Катаболізм
азотистих основ (урацилу, цитозину,
тиміну) полягає в розриві піримідинових
циклів з утворенням у якості продуктів
похідних амінокислот - ^-аланіну та
$-аміноізобутирату. В свою чергу, ^-аланін
розщеплюється до двоокису вуглецю та
аміаку, тоді як $-аміноізобутират може
метаболізуватися подібно до інших
озщеплення
піримідинових нуклеотидів Початкові
етапи катаболізму піримідинових
нуклеотидів, як і в разі пуринових
нуклеотидів, полягають у відщепленні
рибозофосфату з подальшим окисленням
утворених піримідинів. Катаболізм
азотистих основ (урацилу, цитозину,
тиміну) полягає в розриві піримідинових
циклів з утворенням у якості продуктів
похідних амінокислот - ^-аланіну та
$-аміноізобутирату. В свою чергу, ^-аланін
розщеплюється до двоокису вуглецю та
аміаку, тоді як $-аміноізобутират може
метаболізуватися подібно до інших
розгалужених амінокислот з утворенням сукциніл-КоА.
Білет 14
-
Порушення гомеостазу кальцію
Найбільш поширеними клінічно окресленими порушеннями гомеостазу кальцію є патологічні синдроми, пов'язані з дефіцитом вітаміну D3 (які проявляються як рахіт у дітей і різні форми остеопорозу в дорослому та похилому віці) та захворювання, спричинені первинною патологією паращитовидних залоз—гіпер- та гіпопаратиреоз.
Рахіт - захворювання дитячого віку, яке спричиняється зменшеним надходженням та (або) синтезом в організмі вітамінів групи D — тваринного D3 (холекальциферолу) та рослинного D2 (ергокальциферолу). Вірогідність захворювання значно зростає в умовах недостатнього опромінення шкіри дитини сонячним світлом, що є необхідним для утворення вітаміну D3 з 7-дегідрохолестерину. Основними проявами рахіту є гіпо- кальціємія та гіпофосфатемія, які призводять до глибоких порушень кальцифікації кісткової тканини та специфічних змін скелета.
Гіперпаратиреоз - група захворювань, розвиток яких пов'язаний із надлишковою секрецією паратгормону, аномальним збільшенням внаслідок цього концентрації кальцію в сироватці крові і гіпофосфатемією. Первинний гіперпаратиреоз - патологія, що спричиняється наявністю в паращитовидних залозах гормонально активних пухлин - аденоми, карциноми - або гіперплазією залози. Провідними симптомами захворювань цієї групи є ураження кісткової системи (проявляється демінералізацією кісток остеопорозом) та нирок із розвитком сечокам'яної хвороби (внаслідок відкладання солей та утворення каменів, що складаються з оксалату та фосфату кальцію) і нефрокальцинозу.
-
Хімічний склад поперечно-смугастих мязів.
Біохімічний склад м'язів Скелетні м'язи ссавців містять у своєму складі: 72-80 % води; 16-20 % білків; 0,9-2,2 % небілкових азо-і А ! тистих сполук (креатин, креатинфосфат, АТФ,
АДФ, амінокислоти тощо); безазотисті органічні сполуки (глікоген - 0,3- 3,0 %; фосфоліпіди - 0,4— 1,0%; холестерин - 0,06- 0,2 %); мінеральні елементи (К+, Са2+, Na+ тощо).
Білки м'язів Білки скелетних м'язів складаються з водонероз- чинних білків міофібрил (які становлять 75-80 % загального вмісту білків м'язів) і водорозчинних білків саркоплазми - так званого "міогену" До складу фракції міогену входять переважно ферменти, що каталізують ката-
болічні реакції, які забезпечують біоенергетику м'язового скорочення - глікогенфос-
форилаза, ферменти гліколізу, креатинфосфокіназа, аденілаткіназа, кисеньдепо-
нуючий білок міоглобін. У саркосомах містяться ферменти циклу трикарбонових
кислот, біологічного окислення й окисного фосфоритування.
Білки міофібрил До складу міофібрил входять такі білки: 1) до складу товстих ниток - білок міозин; 2) до складу тонких ниток - білки актин, тропоміозин, тропоніновий комплекс {тропонін Т, тропонін І, тропонін Q; 3) білок a-актинін - компонент Z-лінії саркомерів; з цим білком сполучені кінці F-актинових молекул тонких філаментів. Міозин - фібрилярний білок, що утворює товсті філаменти міофібрил. Молекула міозину асиметрична, складається з двох важких поліпептидних ланцюгів, що мають конформацію ос-спіралі й закручені один відносно одного; довжина молекули - 160 нм. N-кінці важких ланцюгів утворюють глобулярні "головки", які нековалентни- ми зв'язками сполучені з додатковими чотирма легкими поліпептидними ланцюгами. В умовах триптичного гідролізу міозин розщеплюється на два фрагменти - меро- міозини: легкий мероміозин — LMM {light meromyosin - англ.) та важкий мероміозин - НММ {heavy meromyosin — англ.). Подальший гідроліз НММ папаїном спричиняє утворення двох ідентичних глобулярних субфрагментів S1 (головок міо зину) і паличкоподібного субфрагмента S2
(рис. 33.3). До складу головок S1 входять каталітичні центри з АТФ-азною активністю і
центри для зв'язування з актином (при відсутності АТФ). Фібрилярні "хвости" молекул міозину
контактують між собою в поздовжньому напрямку, утворюючи товсті філаменти саркомерів, до складу кожного з яких входять близько 400 молекул міозину. Глобулярні головки виступають із зовнішньої поверхні філамента (рис. 33.4). Актин - білок, що існує у двох формах: G- та F-актин. G-актин - глобулярний білок, що має вигляд кулястих молекул діаметром близько 5 нм. Молекули G-актину (субоди- ниці) нековалентно сполучаються між собою, утворюючи намистоподібні утворення -ланцюги фібрилярного F-актину. У м'язових клітинах F-актин представлений фібрилярними структурами, що складаються з двох ланцюгів, переплетених один навколо одного (рис. 33.5). F-актин складає основу будови тонких ниток саркомерів. У складі тонких ниток ланцюги F-актину сполучені з тропоміози- ном і тропонінами. Тропоміозин - білкові молекули витягнутої форми, що складаються з двох поліпептидних ланцюгів (а та 0), які утворю ють подвійну спіраль. Паличкоподібні молекули тропоміозину (довжиною 40 нм і товщиною 2 нм) розміщуються в борозенках між двома ланцюгами F-актину таким
чином, що кожна молекула тропоміозину контактує із сімома молекулами (субоди-
ницями) G-актину. Тропонін - білок тонких філаментів, що складається з трьох субодиниць: ТпТ, ТпІ, ТпС. Тропонінові комплекси мають глобулярну форму і розміщуються впродовж ак-
тинового філамента з інтервалами в 38,5 нм, контактуючи з кінцями молекул
тропоміозину (рис. 33.6). Найбільш вивченим компонентом тропонінового комплексу є ТпС - кальційзв 'язуючий білок, близький за структурою і властивостями до кальмодуліну - універсального трансдуктора кальцієвих сигналів у біохімічних системах. Білок ТпІ взаємодіє з актином, ТпТ - забезпечує взаємодію тропонінового комплексу з тропоміозином.