

Болдырев А.А., Кяйвяряйнен Е.И., Илюха В.А.
БИОМЕМБРАНОЛОГИЯ
Учебноепособие длястудентоввысшихучебныхзаведений,
специализирующихсявобластибиологии, медициныипсихологии
Петрозаводск
2006
УДК 571.1
Болдырев А.А., Кяйвяряйнен Е.И., Илюха В.А. Биомембранология:
Учебное пособие.– Петрозаводск: Изд-во Кар НЦ РАН, 2006.– 226 с., 78 рис., 12 табл.
Учебное пособие «Биомембранология» описывает основные закономерности строения и функционирования клеточных мембран. Книга написана на основании анализа современных достижений клеточной биологии, нейрохимии и иммунологии, во многом отражает экспериментальный опыт самих авторов, много лет работающих в этой области естествознания. Книга подводит итог чтения курса по этой специальности в МГУ им. М.В. Ломоносова, Петрозаводском и Тюменском государственных университетах, а также в Университете штата Нью-Йорк (США). Рекомендуется для студентов и аспирантов естественно-научных вузов и институтов, специализирующихся в области биохимии, органической химии, биотехнологии, физиологии и психологии, а также для специалистов, изучающих широкий круг биологических явлений.
Рецензенты Член.-корр. РАМН, доктор мед. наук, профессор З.А. СУСЛИНА
Доктор биол. наук, профессор Н.Н. НЕМОВА
Научный редактор – профессор С.А. ЧЕПУРНОВ
Издание осуществлено при финансовой поддержке Программы Президиума РАН «Поддержка молодых ученых» (ЭБ НОЦ ИБ КарНЦ РАН) и грантов Президента РФ «Ведущие научные школы» НШ.894.2003.4 и НШ.1760.2003.4
ISBN 5-9274-206-2
©А.А. Болдырев, Е.И. Кяйвяряйнен, В.А. Илюха, 2006
©Институт биологии КарНЦ РАН, 2006
К ЧИТАТЕЛЮ
Когда Эрла Сазерлэнда, первого Нобелевского лауреата в области механизмов клеточной сигнализации, спросили, что больше всего помешало его открытию механизмов передачи сигнала через клеточную мембрану, он ответил:
– Незнание того, как она устроена.
Сегодня для клеточного биолога (нейрохимика, физиолога, биотехнолога, психолога) – незнание законов, по которым функционирует клетка, незнание того, как устроена клеточная мембрана, - нонсенс, несовместимый с выбором перспектив современных исследований в области биологии клетки.
Вот почему мы надеемся, что при всей ограниченности объема этой книги, недостаточной глубине, несовершенстве изложения материала, эта книга найдет своего читателя, как нашел своего слушателя курс «Биомембранология», читаемый в ведущих вузах нашей страны, что, кстати, отличает нашу отечественную систему естественно-научного образования от стран Болонской системы, где этот курс подменяется отрывочными сведениями, излагаемыми «по ходу дела» в лекциях по биохимии, физиологии, биотехнологии или биологии клетки.
Старый спор между преподавателями, что важнее для студента – систематичность и глубина знаний или открытие широких перспектив в их накоплении, мы отважно и безоговорочно решаем в пользу второго фактора: чтобы пытаться найти свое место в науке, надо видеть горизонты ее развития. А когда Вы найдете это место – углубление научных знаний будет Вашей собственной профессией. Мы от души надеемся, что какое бы направление науки Вы не избрали, биомембранология будет одной из основ Вашего естественнонаучного мировоззрения.
Авторы
3
СПИСОК СОКРАЩЕНИЙ
Основные сокращения, принятые в тексте, соответствуют рекомендациям комиссии по биохимической номенклатуре IUPAC.
Другие используемые сокращения приведены ниже:
АФК – активные формы кислорода АЦ – аденилатциклаза ДАГ – диацилглицерол
ДЦКД – дициклогексилкарбодиимида ДЭС – диэтилстильбестрол ИФ3 – инозитол-1,4,5-трисфосфат
ККМ – критическая концентрация мицеллообразования ЛНП – липопротеины низкой плотности ЛОНП – липопротеины очень низкой плотности
ЛПП – липопротеины промежуточной плотности МДА – малоновый диальдегид мХР – мускариновый холинергический рецептор
ПК – протеинкиназа ПОЛ – перекисное окисление липидов СМ – сфингомиелин
СОД – супероксиддисмутаза СР – саркоплазматический ретикулум ТК – тирозинкиназа ФДЭ – фосфодиэстераза
ФИФ2 – фосфатидилинозитол-4,5-бисфосфат ФРК – фактор роста клеток ФРЭ – фактор роста эпидермиса ФХ – фосфатидилхолин ФЭ – фосфатидилэтаноламин ФС – фосфатидилсерин
ЭПР – электронный парамагнитный резонанс ЭР – эндоплазматического ретикулум ЯМР – ядерный магнитный резонанс
GPCR – рецепторы, связанные с G-белками
NANA – N-ацетилнейраминовая кислота РРаза – пирофосфатаза
4
Мембраны играют ключевую роль в структурной организации всех клеток – прокариотических и эукариотических, растительных и животных.
По мере развития биологии определяющая роль мембран в жизни клетки становится все более очевидной. В силу особенностей своего строения мембраны во многом определяют особенности функционирования клеток.
1. ЭВОЛЮЦИЯ ПРЕДСТАВЛЕНИЙ О СТРОЕНИИ МЕМБРАН
Наличие мембран вокруг живых клеток было установлено более ста лет назад в работах К. Негели, который в 1855 году обнаружил, что неповрежденные клетки могут изменять свой объем при изменении осмотического давления окружающей среды. Эти исследования были продолжены Е. Овертоном, показавшим, что неполярные молекулы легче проходят через клеточную мембрану, чем полярные соединения. На основе этих наблюдений он впервые высказал предположение, что клеточная мембрана имеет липидную природу. Развитие идей о структуре мембран существенно продвинулось благодаря работам Е.Гортера и Ф.Грендела, проведенным в 1925 г. Эти авторы впервые выдвинули концепцию липидного бислоя. Эта идея возникла на основе простого эксперимента. Липиды эритроцитов экстрагировали ацетоном и затем получали из них тонкую пленку на поверхности воды. С помощью поплавка сжимали слой липидных молекул на границе раздела вода/воздух до тех пор, пока этот слой не начинал оказывать сопротивление дальнейшему сжатию; это явление было объяснено образованием плотно упакованной мономолекулярной липидной пленки. Измерение площади, занимаемой липидами, и сравнение ее с площадью поверхности эритроцитов, из которых эти липиды были экстрагированы, дали соотношение 2:1. Отсюда был сделан вывод, что мембрана эритроцитов состоит из липидных молекул, расположенных в два слоя. По-видимому, этот вывод Е. Гортера и Ф. Грендела оказался правильным только благодаря взаимной компенсации ошибок (во-первых, экстракция ацетоном извлекает
5
|
не все липиды, во-вторых, они |
|||||
|
дали |
заниженную |
оценку |
|||
|
площади поверхности эритро- |
|||||
|
цитов, использовав для ее |
|||||
|
определения |
высушенные |
||||
|
клетки). Однако в историчес- |
|||||
|
ком плане эта работа имела |
|||||
|
большое значение, |
поскольку |
||||
|
концепция липидного |
бислоя |
||||
|
как |
структурной |
основы |
|||
|
биологических |
мембран |
на |
|||
|
самом деле оказалась верной. |
|||||
|
Мысль о том, что с мембра- |
|||||
|
нами связаны белки, высказана |
|||||
|
десятью годами позже Дж. Да- |
|||||
|
ниелли в связи с необходимо- |
|||||
|
стью объяснить явное расхож- |
|||||
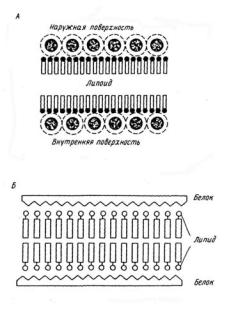
Рис. 1. Модель строения |
дение |
между |
поверхностным |
|||
биологических мембран Даниели- |
натяжением на |
границах |
раз- |
|||
Девсона |
дела |
масло/вода |
и |
мембра- |
||
|
на/вода. Была высказана гипо- |
|||||
|
теза, что мембрана состоит из |
|||||
|
||||||
двойного липидного слоя, и предположено, что белок располагается на ее поверхности – модель Даниели – Дэвсона, или модель «сэндвича» (рис. 1 А, Б). Это была очень удачная модель, и в течение последующих 30 лет многочисленные экспериментальные данные, особенно полученные с помощью дифракции рентгеновских лучей и электронной микроскопии, полностью подтвердили ее адекватность. Основными компонентами биологической мембраны являются липид и белок, вопрос о взаимном расположении этих компонентов в мембране стал предметом многочисленных дискуссий, так как обнаружилось, что мембраны выполняют разнообразные функции.
6
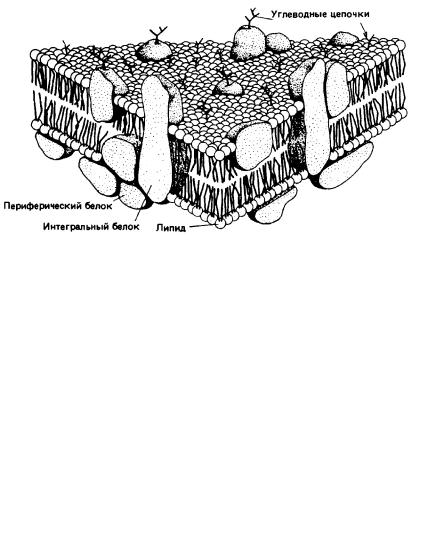
Рис. 2. Жидкостно-мозаичная модель строения биологических мембран
Быстрый прогресс в мембранологии, в результате которого сформировались современные представления, был достигнут в значительной мере благодаря успехам в изучении свойств мембранных белков. Электронно-микроскопические исследования с применением метода замораживания-скалывания показали, что в мембраны встроены глобулярные частицы. Тем временем биохимикам с помощью детергентов удалось «раздробить» мембраны до состояния функционально активных «частиц». Данные спектральных исследований указывали, что для мембранных белков характерно высокое содержание α-спиралей и что они, вероятно, образуют глобулы, а не распределены в виде монослоя на поверхности липидного бислоя. Неполярные свойства мембранных белков наводили на мысль о наличии гидрофобных контактов между белками и внутренней неполярной областью липидного бислоя. С. Сингер и Дж. Никольсон свели воедино все эти идеи, создав жидкост- но-мозаичную модель. В рамках этой модели мембрана представляется как фосфолипидный бислой, в который погружены свободно диффундирующие белки (рис. 2).
7
Прежняя модель Даниели – Дэвсона была статичной и успешно объясняла имевшиеся в то время структурные данные, полученные с довольно низким разрешением. Начиная с 70-х гг. ХХ в. большое внимание стало уделяться изучению динамических свойств мембран и их взаимосвязи с мембранными функциями. В последние годы жидкостно-мозаичная модель (рис.2) также подверглась модификации, и этот процесс будет продолжаться в соответствии совершенствованием наших знаний. Выявляются новые функции цитоскелета. Становится ясно, что не все мембранные белки свободно диффундируют в жидком липидном бислое. Имеются данные о существовании в мембране липидных доменов. Обнаружены динамические ассоциаты липидов, обладающие более плотной упаковкой (рафты). Выявлен специфический класс амфифильных белков, которые под влиянием внеклеточных сигналов меняют свою гидрофобность и обратимо диссоциируют от мембраны. Таким образом, клеточная мембрана все более отличается по своим свойствам от «классического» липидного бислоя. Тем не менее, жидкостномозаичная модель в ее разных модификациях все еще служит в качестве концептуальной основы для объяснения многих мембранных феноменов. Сложность создания единой модели биологических мембран связана с огромным разнообразием мембранных функций.
8
2. КЛЕТОЧНЫЕ МЕМБРАННЫЕ СТРУКТУРЫ
Успехи в исследовании мембран во многом достигнуты благодаря сравнительному изучению мембран из множества разнообразных организмов. Данные, полученные при изучении клеток млекопитающих методом электронной микроскопии, свидетельствуют о наличии развитой сети внутриклеточных мембранных образований (рис. 3). Не вызывает сомнений, что основные принципы структурной организации всех мембран животной клетки по сути одинаковы. Более того, эти принципы распространяются и на мембраны растительных и бактериальных клеток.
Любая клетка имеет наружную мембрану – ее называют плазматической. Она играет роль преграды, отделяющей живое содержимое от ее неживого окружения. Но плазматическая мембрана – не просто оболочка. Она регулирует поступление молекул и ионов в клетку и выход их наружу. Кроме того, в ней находятся различные ферменты, природа которых зависит от особенностей данной клетки. Она содержит специализированные компоненты, участвующие в межклеточных контактах и взаимодействиях, в гормональном ответе и системах транспорта через мембрану как малых, так и больших молекул. Плазматическая мембрана чрезвычайно эластична, благодаря чему животные клетки могут довольно сильно изменять форму без разрыва мембран.
Большинство растительных клеток в отличие от животных не способны изменять свою форму, так как их мембраны окружены толстой, прочной и мало упругой оболочкой. Ее называют клеточной стенкой. Стенки имеются также у бактерий. Бактериальные клетки имеют довольно простую наружную оболочку, содержащую один или два слоя.
Внутри клетки мембраны могут образовывать субклеточные частицы (органеллы) различного назначения. Заметим, что внешний вид органелл неодинаков в клетках разного типа.
9
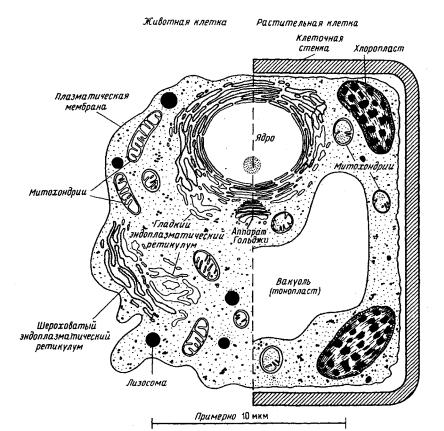
Рис. 3. Схематическое изображение органелл эукариотических клеток животных и растений на основании данных электронной микроскопии
К ним относятся, например, митохондрии - своеобразные энергетические станции клетки, специализированные на образовании АТФ. В этих органеллах осуществляются окислительные превращения субстратов, завершающиеся образованием ATФ. В одной клетке может содержаться от нескольких десятков до нескольких тысяч митохондрий. Они сильно отличаются по размерам и очертанию. Чаще всего митохондрии имеют вид нитей или гранул (по-
10