

pylnev_v_v_red_chastnaya_selekciya_polevyh_kultur
.pdfское 20 (Оренбургский НИИСХ), Барнаульское 98 (Алтайский
НИИ земледелия и селекции сельскохозяйственных культур˝). Эти сорта высокоурожайны, устойчивы к полеганию, осыпа-
нию, засухе, головне, меньше поражаются меланозом, обладаю˝т высоким качеством зерна. Некоторые из них уникальны по св˝оим
свойствам. Квартет — первый отечественный сорт, который˝ наряду с высокой урожайностью и качеством зерна обладает нара˝стаю-
щей комплексной устойчивостью к природным популяциям го˝- ловни различного расового состава. Он не требует предпосевной
обработки семян пестицидами, что позволяет получать экол˝оги-
чески чистую продукцию. Путем сложной ступенчатой гибрид˝изации создан ценный сорт Заряна с очень крупным тонкопленча˝тым зерном (масса 1000 зерен 8,9...9,9 г), высокими технологическими
и кулинарными качествами. Сорт зерноукосного типа Барнау˝ль-
ское 98 удачно сочетает признаки зерновых и кормовых сорто˝в. Кроме сортов крупяного направления в Государственный ре˝-
естр РФ включено три сорта зеленоукосного типа: Абаканско˝е кормовое, Кормовое 45 и Казанское кормовое.
Вопросы для самоконтроля
1.Как меняется характер цветения и опыления проса в зависи˝мости от погодных условий?
2.Каковы основные задачи селекции проса при создании сорт˝ов крупяного и кормового направления?
3.Почему в селекции проса позже, чем у других культур, начал˝и применять метод искусственной гибридизации?
4.Каковы особенности наиболее часто применяемых методов ˝кастрации и опыления проса?
5.Какие типы скрещиваний распространены в селекции проса˝ и с какой целью их применяют?
6.По каким признакам можно эффективно проводить массовый˝ отбор у про-
ñà?
7.Каковы основные показатели технологических качеств про˝са?
231

à ë à â à 11
ГОРОХ
∙
Горох — основная зерновая бобовая культура в Российско˝й Федерации. Он один из главных источников растительного (пищ˝евого и кормового) белка, содержание которого в зерне гороха сос˝тавляет
в среднем 20...27 %. При этом белок гороха содержит в достаточно˝м количестве все 8 незаменимых аминокислот, а содержание ли˝зина в
нем в 1,5...2 раза больше, чем в белке зерновых. По коэффициен-
ту переваримости белок гороха близок к белку куриного яйц˝а и молока.
Поскольку горох был, есть и будет главной зернобобовой ку˝льтурой в России, ВНИИЗБК с соисполнителями (ВНИИ крахмалопродуктов, НИИ новых пищевых веществ) разработали техн˝о-
логию получения продуктов глубокой переработки зерна го˝роха.
Результаты испытания белково-углеводных продуктов, полу˝ченных из зерна гороха, показали, что они могут быть хорошими з˝а-
менителями дорогостоящего импортного соевого концентра˝та при изготовлении колбасных, кондитерских изделий и плавлено˝го сыра,
что делает эти продукты значительно дешевле без снижения˝ питательной ценности и вкусовых качеств.
На нужды животноводства в настоящее время расходуется пр˝и-
мерно 28 млн т зерна, где доля зернобобовых составляет все˝го 5 % вместо необходимых 15...20 %. Дефицит белка в кормах составляет
20...30 г/корм. ед., в результате перерасход кормов на производ˝ство
единицы животноводческой продукции достигает 300...400 % и боле˝е.
Горох — азотфиксирующая культура и играет важную агрот˝ехническую роль. Это один из лучших предшественников почти ˝для всех культур, существенный элемент биологизации земледе˝лия. В южных регионах страны его можно выращивать как промежуто˝ч- ную культуру, что позволяет получать два урожая в год с одной и той же площади.
За рубежом на больших площадях горох возделывают в Китае,˝
Индии, из европейских стран — в Нидерландах, Германии, Ан˝г-
лии, Польше и др.
11.1. СИСТЕМАТИКА И ПРОИСХОЖДЕНИЕ
Горох (род Pisum L.) относится к семейству бобовых (Fabaceae
Lindl., или Leguminosae Juss.), колену виковых (Vicieae Bron.) и включает, по классификации Р. Х. Макашевой, всего два вида˝.
232
Горох красно-желтый (P. fulvum Sibth. еt Smith)— однолетнее
низкорослое растение высотой 10...75 см. Бобы мелкие (3...4 × × 0,7...1 см), прямые. Семена округлые, диаметром 0,3...0,4 см. Встречается в дикой флоре в горных районах Малой Азии и Ар˝а- вии. При скрещивании с горохом посевным обнаруживает высо˝кую
стерильность.
Горох посевной (P. sativum L. sensu amplissimo Govorov) — îäíî-
летний чрезвычайно полиморфный вид, включающий и зимующие формы. Диких форм немного. В сущности, это вид-космопо-
лит. По современной классификации включает шесть подвидо˝в:
высокий (elatius), сирийский (syriacum), абиссинский (abyssinicum), закавказский (transcaucasicum), азиатский (asiaticum) и посевной (sativum).
Горох — древний и типичный род Старого Света, который про˝-
исходит из горных районов Переднеазиатского, Среднеазиа˝тского
èАбиссинского центров (по классификации Н. И. Вавилова).
À.И. Говоров выделил 18 эколого-географических групп в пре˝- делах трех подвидов гороха посевного. Наиболее существен˝ные из
них следующие.
С р е д и з е м н о м о р с к а я — растения очень крупные. Сте-
бель толстый, высотой 80...300 см. Бобы крупные, длиной более 6 см. Масса 1000 семян 250...400 г. Растения отличаются довольно
быстрым темпом роста в период всходы — цветение, требова˝тельны к влаге и относительно устойчивы к понижению температу˝ры
в это время. В последующие фазы устойчивы к периодической˝ за-
сухе, но требовательны к теплу в период налива зерна. В цело˝м вегетационный период средней продолжительности. В оптимальных условиях дают высокие урожаи зерна хорошего качества˝ (со-
держание белка 23...26 %). Сорта: Уладовский 303, Чишминский
ранний и др.
З а п а д н о е в р о п е й с к а я — растения крупные или среднего размера. Высота стебля 40...150 см. Бобы крупные или среднекрупные. Масса 1000 семян 180...300 г. Растения характеризуются медленным ростом и развитием в начале вегетации, но перио˝д цветение—созревание проходит быстро. Более требователь˝ны к влаге, теплу и агрофону, чем растения предыдущей группы. Со˝рта: Роман и Орлик (бывшая Чехословакия), Паули (Нидерланды). К
этой группе относятся главным образом овощные формы.
С р е д н е е в р о п е й с к а я — самая многочисленная группа.
Вегетативные органы средних или, реже, крупных размеров, г˝ене-
ративные — мелких или средних. Масса 1000 семян 150...260 г. Ра-
стения в основном среднерослые, реже высокие. Характеризу˝ются быстрым темпом роста, малотребовательны к влаге в первый ˝период вегетации. Относительно малотребовательны к теплу в˝ тече- ние всего вегетационного периода, высокопластичны, но вме˝сте с тем при интенсивном агрофоне по урожайности могут уступа˝ть
двум предыдущим группам. Имеются ультраскороспелые сорт˝а.
233
Содержание белка в зерне колеблется от 21 до 31 %. Сорта: Капи-
тал, Рамонский 77, Урожайный, Горьковский 186 и др.
С и б и р с к а я — растения средних размеров. Высота стебля до 150 см. Семена мелкие, масса 1000 семян до 200 г. Растения в период всходы—цветение растут медленно, благодаря чему в эт˝о вре-
мя устойчивы к засухе. Менее требовательны к теплу в перио˝д налива и созревания семян. Чувствительны к длине дня. При уко˝ро-
ченном световом дне не зацветают. Сорта: Тулунский гибрид, Град амурский.
11.2. МОРФОБИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
Морфологические признаки. Корень стержневой, проникает в
почву на глубину до 1,5 м, боковые корни расположены в основ˝- ном в пахотном слое почвы. На корнях в симбиозе обитают азо˝т-
фиксирующие бактерии из рода Rhizobium.
Стебель обычно неясночетырехгранный, полый. Он может
быть простым (индетерминантный) с равномерным распредел˝е- нием соцветий по всему стеблю высотой от 50 до 300 см; фасции-
рованным — первая модель детерминантности, описанная е˝ще в
XVI в., но не получившая распространения из-за большой склон˝- ности к полеганию; люпиноидным — уникальная детерминан˝тная
форма с многоплоидным апикальным цветоносом (в настоящее˝ время ее интенсивно прорабатывают в селекционных програ˝ммах)
и, наконец, детерминантным (формы с апикальным соцветием):˝ а) московская модель с морщинистыми семенами, б) луганская˝
модель — генетически идентичная московской, но с гладки˝ми семенами. Они еще не получили широкого применения из-за мало˝-
го числа продуктивных узлов (1...2). Пока же наибольшую селек-
ционную ценность представляет самарская модель детерми˝натности, обнаруженная А. Е. Зубовым в 1983 г. Отличительная особе˝н- ность этой модели — постепенная редукция прилистников ˝в зоне плодоношения и наличие рядом с верхними соцветиями сильн˝о- редуцированной почки. Она названа физиологическим типом˝ детерминантности. Преимущество этого типа детерминантнос˝ти — формирование 3...4 продуктивных узлов (рис.11.1).
Лист гороха обычно состоит из черешка, двух-трех пар листо˝ч-
ков и непарного числа усиков (3 или 5, иногда 7). Встречаются
также и другие типы листа: акациевидный, когда лист не имее˝т
усиков, оканчивается непарным листочком; усатый (безлист˝очковый), когда вместо листочков образуются непарноперистые ˝усики;
многократнонепарноперистый, когда главная многократно ˝разветвленная жилка листа оканчивается тремя—пятью очень м˝елкими листочками (рис.11.2.). В настоящее время созданы гетерофильные мутанты, так называемые хамелеоны, у которых на растении развиваются листья различных типов.
234

3 |
4 |
Рис. 11.1. Стебель гороха:
1 — простой; 2 — фасциированный; 3 — люпиноид; 4 — детерминантный (самарская модель)
235
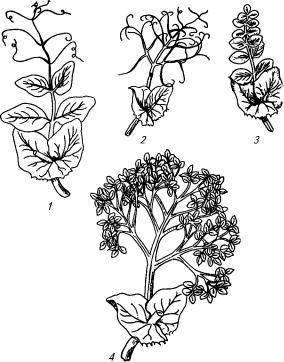
Рис. 11.2. Типы листа гороха:
1 — обычный; 2 — усатый (безлисточковый); 3 — акациевидный (безусиковый); 4 — многократнонепарноперистый
Прилистники у гороха крупнее листочков, полусердцевидно˝й
формы. Однако обнаружены мутанты, у которых прилистники полностью или частично редуцированы. Соцветие — пазушн˝ая кисть, у фасциированных форм — ложный зонтик. Цветок мотылькового типа (рис. 11.3), разной величины и окраски. У сорто˝в зернового или овощного использования окраска белая, корм˝ового и сидерационного — розовая, кармазиновая, красно-фиолет˝овая и т. д. Тычинок 10, одна из них свободная, но тесно прилегает к з˝а-
вязи, остальные девять срастаются, образуя тычиночную тру˝бку.
Завязь почти сидячая, семяпочек 10...12.
Плод — боб — состоит из двух створок, по строению которы˝х различают лущильные и сахарные формы гороха. У первых ств˝ор-
ки боба имеют пергаментный слой и при созревании легко ра˝с- трескиваются, у вторых пергаментного слоя нет и семена плохо обмолачиваются. Бобы могут быть различной формы и размера˝.
236

Рис. 11.3. Части цветка гороха:
1 — парус; 2 — крылья (весла); 3 — лодочка из двух сросшихся лепестков; 4 — завязь со столбиком и тычиночная трубка
Семя состоит из зародыша и се-
менной кожуры. Под ней расположены две семядоли, которые
при прорастании семени остаются в почве. Семена различаются
по крупности, форме и окраске. Рубчик светлый, бурый или чер-
íûé.
Созданы сорта с семенами без рубчика, у которых семяножка срастается с семенной кожурой,
поэтому они не осыпаются
(ðèñ. 11.4).
Биологические особенности. Ãî-
рох — растение умеренного климата. Минимальная температура прорастания 1...2 °С, оптималь˝-
ная — 18...25 °С. Всходы большинства сортов могут переносить˝ кратковременные заморозки до –4...–6 °С.
Нормальная температура для формирования вегетативных о˝р-
ганов 12...16 °С, генеративных — 16...20 °С, для роста бобов и налива семян 16...22 °С. При 25 °С рост замедляется, а при температуре выше 35 °С прекращается.
Оптимальной влажностью для гороха считается 80 % полной
влагоемкости почвы. Критический период в отношении недос˝татка влаги охватывает фазы от начала закладки генеративных орга-
нов до полного цветения. У гороха посевного обнаружены фо˝рмы, относительно устойчивые к засухе в течение всего вегетац˝ионного периода или лишь в определенные
фазы развития.
В целом горох — культура длинного дня, но в пределах вида встре-
чаются формы с разной фотопери-
одической реакцией, которая тесно связана со спектральным составом света. Преобладание длинноволновых лучей ускоряет развитие расте-
|
íèé. |
|
|
Горох — самоопыляющееся рас- |
|
|
тение, однако строение его цветка |
|
Рис. 11.4. Бобы и семена (с разви- |
свидетельствует об энтомофильном |
|
типе опыления в прошлом. Само- |
||
той семяножкой) неосыпающегося |
||
гороха |
опыление, как правило, происхо- |
237
дит в закрытом бутоне, время его наступления зависит от ос˝обен-
ностей сорта, у средиземноморских и европейских групп оно˝ отмечается в момент, когда пестики уже распрямились и напол˝овину вышли из-за зубчиков чашечки, у азиатских форм — в боле˝е ранний период. Прорастание пыльцы начинается примерно че˝рез
15...30 мин, а оплодотворение — через 3...5 ч после нанесения ее на рыльце пестика.
Âусловиях жаркой сухой погоды иногда происходит и перекр˝е- стное опыление, которое составляет доли процента и лишь в˝ ис-
ключительных случаях достигает нескольких процентов. Пр˝едпо-
лагают, что перекрестное опыление осуществляется в основном с помощью личинок трипсов, хотя не исключают участие пчел и˝ шмелей. Установлено, что рыльце сохраняет жизнеспособнос˝ть в
течение 8...10 дней. Однако более высокий процент завязывания˝
семян отмечается при опылении в первые 3 дня после кастрац˝ии.
Âзависимости от сорта и условий выращивания (температур˝ы,
влажности) цветение индетерминантных форм у гороха продо˝лжается 3...40 дней. Более интенсивно цветут фасциированные фор-
мы, люпиноиды, а также формы с детерминантным типом роста.
Âпределах растения первыми зацветают цветки нижнего фер˝тиль-
íîãî óçëà.
11.3. ГЕНЕТИКА
Горох посевной — один из наиболее излюбленных генетиче˝с-
ких объектов. Изучение генетики этой культуры имеет давню˝ю историю и связано еще с именем Т. Найта. Но фундаментальны˝е
работы по генетике гороха были выполнены чешским ученым Г. Менделем. Горох весьма удобен для генетических исследо˝ва-
ний, так как он строгий самоопылитель с довольно контрастными˝
морфологическими признаками и всего семью парами хромосом (2n = 14), т. е. семью группами сцепления.
Согласно обобщению С. Бликста (1972), известно около 1000 мутантных локусов, из которых более 300 идентифицированы, 170 ло-˝ кализованы в семи группах сцепления. Наследование по типу˝ неаллельного взаимодействия, плейотропность многих генов˝ значи- тельно затрудняют генетический анализ наиболее ценных в˝ хозяй-
ственном отношении признаков.
Признаки всходов и стебля. Нормальное развитие хлорофилла
обусловлено присутствием доминантных генов Alb è Àè, их рецес-
сивные аллели вызывают появление соответственно белых и˝ золотистых всходов, которые через 10 дней погибают.
Общая длина стебля гороха детерминируется многими генам˝и. Одни из них контролируют длину междоузлий, другие — их чи˝сло.
Из литературных источников известно, что фасциация стебля — первая модель детерминантности — определяется со˝четанием рецессивных аллелей fa è fas. Растения с генотипами FaFas,
238
Fafas, faFas имеют обычный стебель. Генетика уникальной детер-
минантной формы с многоплодным апикальным цветоносом —˝ люпиноида в настоящее время усиленно изучается. Луганска˝я и московская детерминантность стебля связаны с наличием г˝енаdet.
Он обеспечивает жесткую блокировку апикального роста ст˝ебля:
на апексе отсутствует вегетативная почка и после завязыв˝ания плодов на верхних соцветиях дальнейшее нарастание стебл˝я прак-
тически прекращается. Что касается самарской модели дете˝рминантности, то она контролируется менее жестко геном deh. Расте-
ния с самарским типом детерминантности роста имеют на апе˝ксе
вегетативную (хотя и редуцированную) почку и в определенн˝ых погодных условиях могут возобновлять рост стебля. Коротк˝остебельность ряда сортообразцов обусловлена действием рец˝ессив-
ных генов le è lm, вызывающих уменьшение длины междоузлий.
Åñëè äâà ãåíà Fr è Fru представлены доминантом, ветвление стебля отсутствует, если один из них в рецессиве — Frfru è frFru — ïî-
являются 3...4 ветви, а при frfru образуются 7...10 ветвей. Признаки листьев. Тип листа как главного ассимилирующего
органа существенно влияет на продуктивность растения. Од˝нако важное значение имеют не только площадь листьев, но и их ос˝ве-
щенность, а также поверхностная плотность листьев, обусло˝вливающие количество временно депонированных запасов веще˝ств,
играющих важную роль в формировании семенной продуктивн˝ости, содержание хлорофилла à è b, фотохимическая активность мо-
лекул хлорофилла и т. д.
Растения с акациевидным типом листа имеют большую ассимиляционную поверхность, но из-за сильного полегания эффе˝к- тивность деятельности таких листьев резко снижается всл˝едствие
их взаимозатенения. Растения с усатым типом листа значите˝льно
меньше полегают, но по площади листьев уступают растениям˝ с
обычным типом листа.
По мнению В. В. Хангильдина, тип листа у гороха определяет˝ся сложным взаимодействием доминантных и рецессивных алле˝лей генов Tl è Af (leaf). Òàê, Tl при взаимодействии с Af вызывает развитие обычного типа листа, а при сочетании с af — безлисточкового (усатого) типа листа. Есть и другие мнения.
Многие исследователи считают, что у гороха аналогом флаго˝-
вого листа выступают прицветники — небольшие листовые об˝ра-
зования, расположенные на цветоножке. У некоторых высокоу˝ро-
жайных сортов гороха встречаются растения с небольшими о˝ди-
ночными прицветниками, контролируемыми аллелями генов Br è Bra (BrBrbrabra èëè brbrBraBra). Спонтанный мутант П-1 с крупными парными прицветниками с полной пенетрантностью, кон˝т- ролируемыми рецессивным аллелем brac, обнаружен в посеве сорта гороха Изумруд (А. Н. Зеленов, 2001).
Признаки боба и семян. Большое внимание селекционеров
привлекает такой признак, как развитая семяножка, обуслов˝лен-
239
ный рецессивным геном. Впервые этот признак был обнаружен˝
А. Я. Розенталом и назван tenax. Его современная символика, по В. В. Хангильдину, def (development funiculus). Рецессивное состояние этого гена обеспечивает неосыпаемость семян.
Устойчивость к болезням. Изучение устойчивости гороха к бо-
лезням начато сравнительно давно. Во многих случаях она к˝онтролируется одним, чаще всего доминантным, геном. Так, аллел˝ь
Åï обусловливает устойчивость к вирусу обыкновенной мозаики,
àаллель en — восприимчивость к нему. Гены Fu è Fnu определяют
устойчивость к возбудителям соответственно 1-й и 2-й расы ви˝лта,
àрецессивные аллели fu è fnu — восприимчивость к ним. В отношении мучнистой росы картина обратная: доминантный аллель År обусловливает восприимчивость, а рецессивный аллель år — óñ-
тойчивость к ней. Есть данные, что устойчивость к аскохито˝зу
контролируется тремя доминантными генами.
11.4.ЗАДАЧИ И НАПРАВЛЕНИЯ СЕЛЕКЦИИ
Âиспользовании гороха как полевой культуры, а следовател˝ь-
но, и в его селекции различают три основных направления: пр˝о-
довольственное, зернофуражное и укосно-кормовое.
К сортам всех направлений предъявляются общие требовани˝я:
высокая и устойчивая урожайность при хорошем качестве пр˝одукции, устойчивость к наиболее распространенным болезням (˝аско-
хитозу, антракнозу, мучнистой росе, фузариозным болезням,˝ бактериозу) и вредителям (тле, брухусу и т. д.), устойчивость к р˝ас-
трескиванию бобов и осыпанию семян. С целью создания неос˝ы- пающихся сортов в нашей стране была принята специальная
программа под названием «Тенакс» («Теnах»). Она предусматр˝и-
вала обязательное включение в гибридизацию в качестве од˝ного из компонентов форм с геном устойчивости к осыпанию (def). Подавляющее большинство сортов гороха, включенных в Госр˝е- естр, обладает этим геном.
До недавнего времени вся селекционная работа с горохом ба˝зировалась на традиционном высокостебельном листочковом ˝морфотипе, его потенциальная урожайность была низкой, а реал˝ьная
из-за сильного полегания посевов оставалась на уровне 0,15...0,25 т/га.
Полегаемость стебля была существенным препятствием для˝ широ-
кого возделывания гороха, поскольку затрудняла уборку, не˝ по-
зволяла растениям полностью реализовать биологический ˝потенциал продуктивности.
Ретроспективный анализ стародавних и современных сорто˝в гороха, проведенный во ВНИИЗБК А. Н. Зеленовым и А. В. Амелиным, показал что почти четырехкратное увеличение урожа˝йности, достигнутое за сто лет научной селекции, произошло в р˝е- зультате повышения уборочного индекса с 20 до 50 % и реутилиз˝а-
240