

pylnev_v_v_red_chastnaya_selekciya_polevyh_kultur
.pdfУЧЕБНИКИ И УЧЕБНЫЕ ПОСОБИЯ ДЛЯ СТУДЕНТОВ ВЫСШИХ УЧЕБНЫХ ЗАВЕДЕНИЙ
∙
ЧАСТНАЯ СЕЛЕКЦИЯ ПОЛЕВЫХ КУЛЬТУР
Под редакцией доктора биологических наук,
профессора В. В. Пыльнева
Допущено Министерством сельского хозяйства Российской Федерации в качестве учебника для студентов высших учебных заведений, обучающихся по специальностям 310200 «Агрономия» и 310600 «Селекция и генетика сельскохозяйственных культур»
МОСКВА «КолосС» 2005
1
ÓÄÊ 633:631.52(075.8) ÁÁÊ 42.1—3ÿ73
×-25
À â ò î ð û: В. В. Пыльнев, Ю. Б. Коновалов, Т. И. Хупацария, О. А. Буко, Е. В. Пыльнева, Л. И. Долгодворова, П. М. Конорев, В. С. Рубец, В. М. Пыльнев, А. Н. Березкин, Л. Л. Березкина
Ðå ä à ê ò î ð И. А. Фролова
Ðе ц е н з е н т доктор биологических наук, профессор Ю. Л. Гужов (ÐÓÄÍ)
Частная селекция полевых культур/В. В. Пыльнев,
Ч-25 Ю. Б. Коновалов, Т. И. Хупацария и др.; Под ред. В. В. Пыль-
нева. — М.: КолосС, 2005. — 552 с.: ил. — (Учебники и учеб. пособия для студентов высш. учеб. заведений).
ISBN 5—9532—0316—0
Рассмотрены основные вопросы селекции важнейших полевы˝х культур: зерновых, зернобобовых, масличных, прядильных, а также кар˝тофеля, сахарной и кормовой свеклы, вики, рапса, нута, многолетних кор˝мовых трав. Изложены систематика и происхождение, морфобиологическ˝ие особенности, генетика, задачи и направления селекции, модели сортов˝ (где они существуют), исходный материал, методы и специальные направле˝ния селекции, методика и техника селекционного процесса, достижени˝я селекции.
Для студентов высших учебных заведений, обучающихся по сп˝ециальностям «Агрономия» и «Селекция и генетика сельскохозяйс˝твенных культур», а также для специалистов в области селекции растени˝й и всех тех, кто интересуется этой проблемой.
ISBN 5—9532—0316—0
ÓÄÊ 633:631.52(075.8) ÁÁÊ 42.1—3ÿ73
©Коллектив авторов, 2005
©Издательство «КолосС», 2005
2

ПРЕДИСЛОВИЕ
∙
Практическая деятельность ученых-агрономов невозможна ˝без глубокого знания ими специальных дисциплин, в частности г˝енетики, селекции и семеноводства. Особенно это касается спе˝циа-
листов в области селекции и генетики сельскохозяйственн˝ых культур. Планомерная подготовка таких специалистов закл˝ючает-
ся в последовательном освоении слушателями основ генети˝ки, об-
щей селекции и семеноводства сельскохозяйственных раст˝ений. Однако их подготовка была бы неполной без специальных зна˝ний
в области селекции отдельных культур, т. е. частной селек˝ции. Естественно, что селекционная работа с отдельными культура˝ми имеет свои особенности, связанные с их биологией. При этом˝ не-
обходимо знание особенностей генетики, селекционной тех˝ноло-
гии, доноров и источников селекционно ценных свойств и пр˝и- знаков отдельных культур.
Каждая глава учебника посвящена селекции отдельной поле˝- вой культуры. Содержание каждой главы максимально унифиц˝и-
ровано. Рассмотрены хозяйственное значение культуры, ее о˝б- щепринятая систематика и происхождение, морфобиологиче˝с-
кие особенности. Особое внимание уделено генетике культу˝ры,
указаны основные селекционно ценные гены отдельных приз˝наков и свойств, характер наследования морфологических при˝зна-
ков при гибридизации. Приведены общие и специфичные на-
правления селекции данной культуры. По возможности даны м˝о-
дели сортов для зон наибольшего распространения культур˝ы с учетом направления ее использования. Широко представлен˝ исходный материал для селекции определенной культуры с ука˝занием отдельных сортов и линий — доноров и источников опр˝е- деленных ценных признаков и свойств, генов, которые несут˝ те или иные источники. Описаны как основные, ставшие традици˝-
онными методы создания новых сортов и гибридов (внутриви˝до-
вая и отдаленная гибридизация, мутагенез, полиплоидия), та˝к и
специфичные для определенной культуры (использование хр˝о-
мосомной инженерии, биотехнологии, ядерной и мужской сте-˝ рильности и т. д.). При раскрытии методики и техники селекц˝и- онного процесса каждой культуры особое внимание уделено˝ се-
лекционным оценкам специфических для нее признаков и
свойств. Приведены важнейшие достижения в области селекц˝ии данной культуры с указанием основных селекционных учреж˝дений и ведущих селекционеров. Каждая глава содержит ряд во˝п-
3
росов, охватывающих основные положения изложенного в гла˝ве
материала.
Естественно, что досконально знать селекцию отдельной се˝льскохозяйственной культуры или группы культур может толь˝ко че- ловек, на практике занимающийся данной работой. Поэтому
учебник написан довольно большим авторским коллективом˝, представленным в основном преподавателями Московской с˝ель-
скохозяйственной академии им. К. А. Тимирязева.
Отдельные главы написали: предисловие, «Овес» — професс˝ор
В. В. Пыльнев; «Пшеница», «Ячмень» — профессора Ю. Б. Кон˝о-
валов и В. В. Пыльнев; «Рожь», «Рис», «Гречиха», «Просо» — д˝о-
öåíò Î. À. Áóêî; |
«Тритикале» — профессор В. М. Пыльнев |
(Одесский аграрный |
университет, Украина); «Кукуруза» — про- |
фессор В. В. Пыльнев и доцент Т. И. Хупацария; «Сорго», «Го-˝
рох», «Вика» — доцент |
Т. И. Хупацария; |
«Сахарная |
и кормов˝ая |
||
свекла» |
— профессор |
А. Н. Березкин; |
«Фасоль», |
«Чечевица», |
|
«Ñîÿ», |
«Кормовые бобы», «Нут» — старший |
преподаватель |
|||
Е. В. Пыльнева; «Люпин», «Рапс» — |
старший |
преподаватель |
|||
П. М. Конорев; «Многолетние травы» — доцент Л. И. Долгодворова и старший преподаватель П. М. Конорев; «Подсолнечник» ˝—
профессор В. В. Пыльнев и доцент Л. И. Долгодворова; «Лен-дол- гунец», «Конопля» — профессор Ю. Б. Коновалов; «Картофел˝ь» —
ведущий научный сотрудник Л. Л. Березкина; «Горчица сарептская», «Клещевина» — старший преподаватель В. С. Рубец.
Предлагаемый учебник предназначен в первую очередь для
студентов высших учебных заведений, обучающихся по специ˝альностям «Агрономия» и «Селекция и генетика сельскохозяйс˝твенных культур», однако представляет интерес как для специал˝истов
в области селекции растений, так и для всех, интересующихс˝я
данными вопросами.
4

à ë à â à 1
ПШЕНИЦА
∙
Пшеница занимает первое место в мировом земледелии по пос˝евным площадям и валовому сбору зерна. Ее возделывают главн˝ым образом для производства муки, из которой изготавливают хле˝бобулоч-
ные, кондитерские или макаронные изделия. Из зерна пшениц˝ы вырабатывают также различные крупы. Специальные сорта пшен˝ицы
служат для получения зеленого корма. В качестве корма для˝ скота ис-
пользуют также и солому пшеницы. Почти 3/4 населения нашей п˝ланеты питается продуктами, получаемыми из зерна пшеницы.
СНГ занимает первое место в мире по посевным площадям и валовым сборам зерна пшеницы. Примерно 2/3 посевных площадей отводится под яровую пшеницу, 1/3 — под озимую. Основные˝
районы выращивания озимой пшеницы в России — Северный
Кавказ, Центрально-Черноземная зона, яровой — Поволжье, ю˝говосток европейской части страны, Западная Сибирь.
Крупнейшими производителями зерна пшеницы являются так˝- же США, Канада, Аргентина, Австралия. Значительное количе-
ство его производят Мексика, Бразилия, КНР, Индия. Большие площади под этой культурой заняты в Украине, Казахстане, В˝енг-
рии, странах бывшей Югославии, Франции.
В мире в основном возделывают два вида пшеницы: мягкую (Triticum aestivum L.) и твердую (Triticum durum L.). На долю после-
дней приходится около 10 % посевных площадей, занимаемых
этой культурой. Помимо России и Украины твердую пшеницу
возделывают в Италии, Алжире, Сирии, Турции, Ираке, США, Канаде, Мексике, Аргентине.
Пшеница — одна из родоначальниц новой культуры — трити˝- кале, имеющей кормовое и продовольственное значение и пол˝у- чающей все большее распространение.
1.1. СИСТЕМАТИКА И ПРОИСХОЖДЕНИЕ
Род пшеница (Triticum L.) относят к семейству мятликовых (Poaceae Barnhart), или злаковых (Gramineae Juss). Он входит в
трибу пшенициевых (Triticeae Dum.). В «Культурной флоре СССР»
(Л.: Колос, 1979) указано 27 видов пшеницы. Однако в других источниках приведены иные числа, что зависит от критериев, с˝ ко-
торыми различные авторы подходили к выделению видов внут˝ри рода Triticum.
5
По числу хромосом в соматических клетках все виды пшеницы˝
делят на три группы: 42, 28 и 14-хромосомные, а по пленчатости — на пленчатые (зерновка плотно заключена в цветковые˝ че- шуи, колосовой стержень ломкий) и голозерные (зерновка лег˝ко выделяется из цветковых чешуй, колосовой стержень неломк˝ий) .
Филогения рода Triticum детально исследована с помощью морфологического, цитологического, иммунохимического и эле˝ктро-
форетического методов. Наиболее древние — 14-хромосомные˝ пленчатые виды, самые молодые — 42-хромосомные голозерные˝.
Вся система рода Triticum — результат эволюции двух 14-хромо-
сомных видов, близких к ныне существующим Ò. urartu Thum. ex. Gandil. è Ò. boeoticum Boiss., которые, в свою очередь, по-видимо- му, имели общего предка. Выдающуюся роль в эволюции рода
Triticum сыграла аллоплоидия: в результате спонтанных скрещива-
ний пшеницы с 14-хромосомными видами эгилопса появились 28- и 42-хромосомные виды пшеницы.
Первоначальный геном пшеницы (14-хромосомных видов) принято обозначать буквой À. Если он происходит от вида, близ-
êîãî ê Ò. urartu, то обозначается Àu, если от близкого к Ò. boeoticum — òî Àb. Второй геном (28-хромосомных видов) пше-
ницы, ведущих свое происхождение от T. urartu, — Â — привнесен Aegilops longissima Schweinf. et Muschl., а геном тех видов, которые
происходят от Ò. boeoticum, — G — принадлежит A. speltoides Tausch. Наконец, третий геном (42-хромосомных видов) пшениц, род-
ственных Ò. urartu, — D — получен от A. tauschii Coss. (A. squarrosa
Griseb.). Естественная эволюция Ò. boeoticum не достигла трехгеномного уровня.
Все сказанное иллюстрирует схема филогенетических связ˝ей в
ðîäå Triticum (рис. 1.1). В схеме представлены не все виды рода, а
только наиболее значимые в хозяйственном отношении, а так˝же
иллюстрирующие параллелизм в развитии видов, родственны˝х T. urartu è Ò. boeoticum, и видов, принадлежащих к различным группам по числу хромосом. Так, 28-хромосомная группа, родственная Ò. urartu, включает как пленчатые, так и голозерные виды. Тот же факт давно установлен и для 42-хромосомной груп˝- пы. Однако в 28-хромосомной группе видов, родственных Ò. boeoticum, голозерный вид Ò. mulitinae Zhuk. et Mugush. открыт
недавно. Следует отметить, что в 14-хромосомной группе этих ˝ви-
дов пшеницы также обнаружен голозерный вид (Ò. sinskajae
A. Filat. et Кurk.), в то время как в аналогичной группе с геномом
Àu он пока неизвестен. Указанные выше виды возникли как голо-
зерные мутанты соответственно пшеницы Тимофеева (T. timopheevii Zhuk.) и культурной однозернянки (T. monococcum L.). У пшеницы с геномом Àb известен 42-хромосомный пленчатый вид: пшеница Жуковского (Ò. zhukovskyi Menabde et Ericzjan.), но он возник путем удвоения геномов Àb при спонтанной гибридизации культур-
ной однозернянки и Ò. timopheevii. Таким образом, у видов пшени-
6
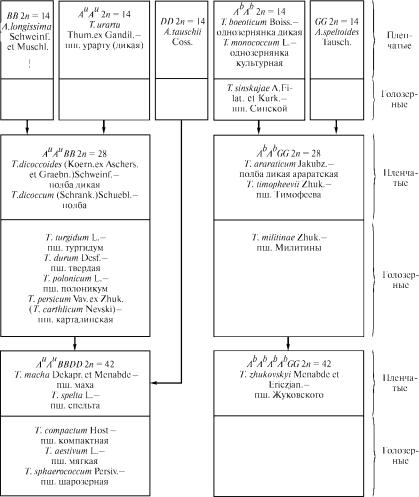
Рис. 1.1. Схема филогенетических связей в роде Triticum L.
цы с геномом Àb нет природного 42-хромосомного аналога пше-
ницы с геномом Àu. Такой аналог создан экспериментально путем
скрещивания Ò. timopheevii è T. tauschii. Это пленчатая пшеница Кихары (Ò. kiharae Dorof. et Migusch.).
Родственные связи внутри отдельных групп видов пшеницы отчетливо не установлены. Так, одни авторы рассматривают ˝пшеницу персидскую, или карталинскую (Ò. persicum Vav. ex Zhuk.,
7
èëè T. carthlicum Nevski), относящуюся к 28-хромосомной группе,
но имеющую морфологическое сходство с пшеницей мягкой ка˝к голозерную мутацию, а другие — как спонтанный гибрид пше˝ницы мягкой с каким-то тетраплоидным видом.
В появлении отдельных видов и целых групп видов выдающую-˝
ся роль сыграли некоторые гены. Так, ген Q обусловливает голозерность у 42-хромосомной группы пшеницы (геном AuBD). Ýòîò
же ген обнаружен у пшеницы карталинской из 28-хромосомной группы (геном ÀuÂ), а его аналог Qm — ó Ò. militinae Zhuk. (геном
AbG). В то же время голозерность других видов данной группы
имеет полигенную природу. Доминантный аллель гена Ñ обусловливает большую плотность колоса, а его рецессивный аллель˝ñ — рыхлость. Основное отличие очень плотноколосой Ò. compactum
Host îò Ò. aestivum L. определяется исключительно различием в
этих аллелях. Аналогично объясняется отличие пшеницы мяг˝кой (удлиненное зерно, аллель s) от шарозерной (аллель S). Доминан-
тный аллель Ð обусловливает длинные колосковые чешуи у
Ò. polonicum L.
Столь незначительные генетические различия и легкая скр˝е- щиваемость видов в пределах групп часто побуждали систем˝ати-
ков объединять родственные виды в один. Наиболее ярко эта тенденция выражена в системе, предложенной шведским генетик˝ом
Дж. Мак-Кеем, который оставил в роде Triticum всего пять видов:
T. monococcum, T. timopheevii, T. zhukovskyi, T. turgidum è T. aestivum
(Mac Key, 1963). Однако представители школы тритикологов Все-
российского научно-исследовательского института растен˝иеводства им. Н. И. Вавилова (ВИР) считают, что различие даже в аллелях одного локуса, если оно меняет хозяйственную характер˝исти-
ку растения (например, пленчатость — голозерность), дост˝аточно
для признания его видовым отличием.
Центр происхождения большинства видов пшеницы — Переднеазиатский генцентр, который включает и Закавказье СНГ. Им˝енно здесь встречаются виды, близкие к предполагаемым родон˝а- чальникам Ò. urartu è Ò. boeoticum, и обнаружено наибольшее разнообразие видов пшеницы, в том числе эндемичных, и множест˝во разновидностей. Однако, по мнению Н. И. Вавилова, родина пш˝е- ниц мягкой, компактной и шарозерной — Среднеазиатский г˝ен-
центр (в Переднеазиатском генцентре имеется эндемичная г˝руппа
пшеницы мягкой). Пшеница твердая возникла как культура па˝-
раллельно в Передней Азии, Эфиопии и Средиземноморье. В
Средиземноморском генцентре и Эфиопии появились также н˝е-
которые другие виды пшеницы. С развитием торгово-экономи-˝ ческих связей пшеница расселилась по всему земному шару. ˝Это прежде всего относится к такому виду-космополиту, как пше˝ница мягкая. Ареал пшеницы твердой примерно очерчен в начале г˝лавы. Среди этих двух видов пшеницы преобладают яровые форм˝ы,
но встречаются озимые и полуозимые. Площади, занятые друг˝ими
8
видами пшеницы, незначительны. Географическое распростр˝ане-
ние наиболее важных из них (в хозяйственном и селекционно˝м отношениях) можно охарактеризовать следующим образом.
Пшеница компактная (карликовая) встречается в Армении (за˝- регистрированные сорта), Турции, Афганистане, Сирии, Китае˝,
Монголии, США. Представлена озимыми, яровыми и полуозимыми формами. Пшеница шарозерная эндемична для полуострова˝
Индостан, причем известны только яровые ее формы. Пшеница˝ тургидум распространена главным образом на Балканах, на ю˝ге и
западе Европы, в Передней Азии, Афганистане, Иране, Восточ-
ном Китае. В СНГ она встречается в Средней Азии, Казахстане и Дагестане. Обнаружены яровые, озимые и полуозимые формы. Пшеницу карталинскую возделывают в Иране и Грузии. Извест˝ны
только яровые формы. Пшеница полоникум сопутствует пшени˝це
твердой, причем известны только яровые ее формы. Полба (дву˝- зернянка, эммер) обнаружена на Балканах, в Средиземноморь˝е,
Эфиопии, Малой Азии, Индии, Закавказье, а в нашей стране — в˝ Среднем Поволжье (Татарстан, Удмуртия, Чувашия). Она пред-
ставлена преимущественно яровыми формами. Спельту можно˝ встретить в горных районах Центральной Европы, на Балкана˝х,
на севере Испании, в Иране, Средней Азии и Закавказье. Найде˝- ны как озимые, так и яровые формы. Культурную однозернянку˝
можно обнаружить на Балканах, в Средиземноморье, горных районах Центральной Европы, Дании, Швеции, Чехии, Слова-
кии, Турции, а в СНГ — в Закавказье, Крыму, на Северном Кав-
казе. Озимые формы очень редки. Пшеницы Тимофеева (яровая), маха (озимая и полуозимая) и Жуковского — эндемики Г˝рузии.
В странах СНГ насчитывают 11 экологических групп пшеницы
мягкой. Среди них довольно много узколокализованных. Наи-˝
большее значение имеют северорусская и сборные группы —˝ степная и лесостепная.
С е в е р о р у с с к а я г р у п п а локализована в Нечерноземной зоне России, западных областях Белоруссии и Украины. Д˝ля нее характерны в основном озимые формы, устойчивые к глуб˝о- кому снежному покрову, влаголюбивые, высокоили среднеро˝с- лые, довольно облиственные и интенсивно кустящиеся.
С т е п н а я г р у п п а в широком смысле включает степную
волжскую, степную южную (украинскую), степную южную (севе-
рокавказскую), степную восточную (Сибирь, Северный Казах-
стан) экологические группы. Относящиеся к ней формы отлич˝а-
ются засухоустойчивостью. Озимые сорта характеризуются˝ высокой морозостойкостью.
Л е с о с т е п н а я г р у п п а также состоит из ряда экологи- ческих групп, локализованных как в европейской части, так˝ и в Сибири. По морозостойкости и засухоустойчивости уступае˝т степ-
ной группе. Для Западной Сибири характерны яровые позднес˝пе-
9
лые сорта с медленным развитием во время кущения и вследс˝твие
этого устойчивые к весенней засухе.
К о р о т к о с т е б е л ь н а я г и б р и д н а я экологическая группа приобретает все большое значение. В нее входят зар˝убежные яровые короткостебельные сорта (из США, Мексики, Ин-
äèè).
Пшеница твердая в СНГ представлена пятью экологическими˝
группами. Наибольшее значение имеет степная группа (в шир˝о- ком смысле), отличающаяся засухоустойчивостью, средней к˝усти-
стостью и облиственностью входящих в нее форм. Представле˝на
яровыми и озимыми сортами.
1.2. МОРФОБИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
Морфология растения пшеницы типична для злака. Корень
мочковатый. Различают зародышевые, колеоптильные и узловые корни. Зародышевые корни появляются при прорастании семени, колеоптильные — из подземного колеоптильного у˝зла.
Вместе они составляют первичную корневую систему, проник˝а-
ющую на большую глубину, но сравнительно мало разветвляю-˝ щуюся в верхних слоях почвы. Узловые корни развиваются из
подземных сближенных узлов главного и боковых побегов и располагаются в верхних слоях почвы. Подземные сближенны˝е
узлы представляют собой зону, или узел, кущения. В пазухах листьев, соответствующих этим узлам, образуются боковые п˝о-
беги. Большое значение имеет глубина залегания узла кущен˝ия. Чем она меньше, тем интенсивнее кущение в условиях доста-
точного увлажнения, но и тем сильнее влияние засухи на это˝т процесс и на развитие узловых корней и тем ниже морозосто˝й-
кость у озимой пшеницы.
Стебель, разделенный на узлы и междоузлия, полый, но может
быть заполнен рыхлой паренхимной тканью. Листья состоят и˝з
влагалища, охватывающего стебель, и линейной листовой пла˝с-
тинки. В месте отгиба листовой пластинки имеется язычок (л˝игу-
ла), защищающий внутреннюю часть влагалища от дождевой во˝ды (встречаются и безъязычковые формы). Нижней частью влагал˝и- ще прикрепляется к основанию стеблевого узла, образуя над˝ ним утолщение — листовой узел.
Соцветие — сложный колос. Оно состоит из колосового стер˝ж- ня и колосков, сидящих на его уступах. У каждого колоска две˝ колосковые чешуи и три-четыре цветка. У пшеницы мягкой число˝ развитых цветков в колоске может достигать пяти-шести, у т˝вердой их меньше. Наиболее развитые колоски формируются несколько ниже середины колоса. В основании колоса обычно р˝асположены один-два недоразвитых колоска, но их может и не бы˝ть. В колоске лучше всего развиты первый и второй цветки, в многоцветковом колоске им не уступает третий. Верхние цветки в˝ ко-
10