

Семинарское занятие № 4 Тема «Физиология деятельности зрительного, слухового и вестибулярного анализаторов»
Вопросы для обсуждения:
1. Общие принципы строения и функции сенсорных систем.
2. Строение и функции оптического аппарата глаза. Рецепторный уровень зрительной системы.
3. Проводящие пути зрительной системы. Зрительная кора.
4. Физические и физиологические параметра звука. Механизмы восприятия звука.
5. Рецепторный уровень и проводящие пути слухового анализатора.
6. Виды слуха. Слуховая кора.
7. Вестибулярная система.
Задания:
По мере изучения сенсорных систем заполните таблицу «Компоненты основных сенсорных систем»:
-
Модальность сенсорной системы
Рецептор
Проводящие пути
Первичные воспринимающие центры коры
Вторичные воспринимающие центры коры
Зрительная
Слуховая
Вестибулярная
Тактильная
Обонятельная
Вкусовая
Проприоцептивная
Висцеральная
Литература:
Психофизиология. Под ред. Ю.И. Александрова. - СПб.: Питер, 2003.
Физиология человека: Compendium. Учебник для высших учебных заведений / Под ред. Акад РАМН Б.И.Ткаченко и проф. В.Ф.Пятина, СПб. – 1996, с. 272 – 277.
Смирнов В.М., Будылина С.М. Физиология сенсорных систем и высшая нервная деятельность: Учеб. пособие для студ. высш. учеб. заведений. – М.: Академия, 2003. – с. 35 – 67.
Хомская Е.Д. Нейропсихология. – СПб.: Питер, 2005 (смотри 8, 10 главы).
Материалы для подготовки к занятию
Физиология человека: Compendium. Учебник для высших учебных заведений / Под ред. Акад РАМН Б.И.Ткаченко и проф. В.Ф.Пятина, СПб. – 1996, 424 с.
Зрительная система
Зрительная система (зрительный анализатор) представляет собой совокупность защитных, оптических, рецепторных и нервных структур, воспринимающих и анализирующих световые раздражители. Свет, как электромагнитное излучение с различными длинами волн - от коротких (красная область спектра) до длинных (синяя область спектра), характеризуется частотой и интенсивностью. Воспринимаемая частота (величина обратная длине волны) определяет окраску света. Интенсивность или яркость имеет диапазон от порога восприятия до болевого порога (160 дБ), т. е. порядка 1016. Через зрительную систему человек получает более 80% информации о внешнем мире за счет пространственной разрешающей способности (острота зрения), временной разрешающей способности (время суммации и критическая частота мельканий), порога чувствительности и адаптации.
Периферический отдел зрительной системы. Глаз - сферическое образование, состоящее из склеры, конъюнктивы, роговицы, радужной оболочки. Склера представляет собой соединительную ткань белого цвета, окружающую глазное яблоко. Конъюнктива - прозрачная ткань, снабженная кровеносными сосудами, которая на переднем полюсе глаза соединяется с роговицей. Роговица является прозрачным образованием, кривизна поверхности которого определяет особенности преломления света. Роговица и конъюнктива покрыты тонкой пленкой слезной жидкости, секретируемой слезными железами, которые расположены в височной части глазницы, над глазным яблоком. Слезы защищают роговицу и конъюнктиву от высыхания. Позади роговицы находится пигментированная радужная оболочка. Между роговицей и радужной оболочкой находится передняя камера глаза, наполненная жидкостью — водянистой влагой. В центре радужной оболочки находится зрачок круглой формы, пропускающий внутрь глаза свет после его прохождения через роговицу.
Размер зрачка в зависимости от освещенности автоматически регулируется нервными волокнами, заканчивающимися в мускулатуре радужной оболочки. Круговая мышца (сфинктер), суживающая зрачок, иннервируется парасимпатическими волокнами, а мышца, расширяющая зрачок (дилататор), иннервируется симпатическими волокнами. Изменения диаметра зрачка меняют интенсивность светового раздражения в 16 – 17 раз. Реакция расширения зрачка до максимального диаметра длится около 5 минут, а максимальное уменьшение его диаметра достигается всего за 5 секунд. Следовательно, основная функция зрачка - это регуляция интенсивности света, который попадает на центральную часть хрусталика, где фокусировка наиболее точная.
Позади радужной оболочки находятся задняя камера глаза и хрусталик. Хрусталик - двояковыпуклая линза» расположен в капсуле, волокна которой соединены с ресничными мышцами и наружным сосудистым слоем сетчатки. Хрусталик может становиться более плоским (рассматривание далеких объектов) или более выпуклым (рассматривание близких объектов). Изменение кривизны хрусталика называется аккомодацией. Функция аккомодации хрусталика - проекция изображения точно на сетчатку. Внутри глаза, позади хрусталика, находится стекловидное тело. Оно представляет собой коллоидный раствор гиалуроновой кислоты во внеклеточной жидкости.
Если расстояние между хрусталиком и сетчаткой больше, чем фокусное расстояние хрусталика, то возникает близорукость (миопия). Если сетчатка слишком близко к хрусталику и фокусировка хороша только при рассматривании далеко расположенных предметов, возникает дальнозоркость (гиперметропия). Близорукость и дальнозоркость корректируются очками с вогнутыми и выпуклыми линзами, соответственно. Изменение лучепреломления в различных диаметрах роговицы называется астигматизмом (результат неравномерной кривизны роговицы). Для его исправления более пригодны контактные линзы, которые, плавая в слезной жидкости над роговицей, компенсируют ее отклонения от правильной формы. Итак, оптическая система глаз фокусирует изображение на рецепторной поверхности сетчатки. Диоптрический аппарат глаза передает на сетчатку резко уменьшенное изображение предметов.
Сетчатка — это высокоорганизованная слоистая структура, объединяющая рецепторы и нейроны. Фоторецепторные клетки (палочки и колбочки) расположены в пигментном слое. Их светочувствительные концы повернуты от пучка падающего света и спрятаны в промежутках между клетками пигментного эпителия. Пигментные клетки сетчатки участвуют в метаболизме фоторецепторов и синтезе зрительных пигментов. Нервные волокна, выходящие из сетчатки, лежат на пути света к ее рецепторам. В том месте, где волокна зрительного нерва выходят из сетчатки, нет фоторецепторов -это так называемое слепое пятно. В области слепого пятна всегда имеется «дефект» изображения, проецируемого на сетчатку, который компенсируют высшие зрительные центры. Поэтому слепое пятно не влияет на целостность зрительного восприятия.
Палочки отличаются от колбочек структурно и функционально. В палочках содержится зрительный пигмент (пурпур — родопсин), а в каждой колбочке зрительные пигменты — иодопсин, хлоролаб и эритлаб, необходимые для цветового зрения. Палочка в 500 раз более чувствительна к свету, чем колбочка, но она не цветочувствительна. Зрительные пигменты расположены в наружном светочувствительном сегменте палочек и колбочек. Во внутреннем сегменте находятся ядро и митохондрии, участвующие в энергетических процессах при действии света. В сетчатке находится около 6 млн. колбочек и 120 млн. палочек. Плотность колбочек выше в центре сетчатки и падает к периферии. В центре сетчатки имеется участок максимальной остроты зрения, называемый центральной ямкой, где находятся только колбочки (150 тысяч на 1 мм2). Палочек больше на периферии сетчатки, но острота «периферического» зрения при хорошей освещенности невелика. Колбочки функционируют при ярком свете и реагируют на цвет, палочки возбуждаются на действие света и обеспечивают зрительное восприятие при слабой (сумеречной) освещенности.
Первичным процессом зрительной рецепции является фотохимическая реакция. Фотоны поглощаются молекулами зрительных пигментов. Поглощение молекулой пигмента одного фотона (кванта света) запускает в фоторецепторе многоступенчатый процесс распада молекул пигмента. При распаде родопсина в палочках, иодопсина, хлоролаба и эритлаба в колбочках образуется опсин и витамин А1 Фотохимические процессы в палочках и колбочках сходны, но каждый пигмент имеет свои спектры поглощения: у родопсина это 500 нм (зелено-голубая часть), иодопсина – 570 нм (желтая часть), хлоролаба – 425 нм, эритлаба – 435 нм. Восстанавливаются пигменты в темноте в результате цепи химических реакций (ресжтез), протекающих с поглощением энергии. Ретиналь ресинтезируется на основе цисизомера витамина А,. Поэтому недостаток витамина А1 в организме вызывает нарушение сумеречного зрения. При постоянном освещении фотохимический распад пигментов уравновешивается с их ресинтезом.
На свет фоторецепторы отвечают гиперполяризацией мембраны (зрительный рецепторный потенциал). Амплитуда этого потенциала прямо зависит
от интенсивности света (освещенности, относительно предыдущего состояния адаптации) и длины его волны.
Палочки и колбочки соединены с биполярными нейронами сетчатки, которые, в свою очередь, имеют с ганглиозными клетками синапсы, выделяющие ацетилхолин. Аксоны ганглиозных клеток сетчатки образуют зрительный нерв, который идет к различным мозговым структурам. Около 130 млн. фоторецепторов связаны (конвергируют) с 1,3 млн. волокон зрительного нерва. В центральной ямке каждая колбочка связана с одной биполярной клеткой, а она, в свою очередь, - с одной ганглиозной. К периферии от центральной ямки множество палочек и колбочек конвергируют на одной биполярной клетке, а множество биполярных — на одной ганглиозной.
Распространение зрительного возбуждения внутри сетчатки ограничивается тормозными нейронами — это горизонтальные и амакриновые клетки, которые расположены в слое биполярных нейронов. Горизонтальные клетки обеспечивают латеральное торможение между биполярными нейронами, а амакриновые - между ганглиозными клетками.
Ганглиозные клетки при слабой освещенности дают непрерывную импульсацию. При большой освещенности одни ганглиозные клетки реагируют как детекторы яркости (усиливают активность), а другие - как детекторы темноты (ослабляют активность). Все ганглиозные клетки имеют круглые рецептивные поля. Оптимальным стимулом для ганглиозных клеток служит либо светлое пятно, окруженное темным поясом, либо темное пятно, окруженное ярким поясом. Многие ганглиозные клетки реагируют только на изменение освещения, но не реагируют на постоянный свет.
Каждая ганглиозная клетка связана со своим рецептивным полем на ограниченном участке сетчатки и не реагирует на свет вне ее рецептивного поля. Причем один тип ганглиозных клеток возбуждается светом, попадающим в центр рецептивного поля, но затормаживается, если свет попадает на его периферию, а другой тип затормаживается светом в центре рецептивного поля и возбуждается при действии света на его края. Одновременно реакция нейронов первого и второго типов лежит в основе появления одновременного контраста за счет подчеркивания края изображения их антагонистическими рецептивными полями.
Величина рецептивных полей растет от области центральной ямки к периферии сетчатки (жцентриситет). Это связано с тем, что в середине сетчатки, в области центральной ямки, одна колбочка через отдельную биполярную клетку соединяется с одной ганглиозной клеткой. На периферии, где, в основном, находятся палочки, отмечаются широкие перекрывающиеся рецептивные поля: множество рецепторов связано с одной ганглиозной клеткой.
Острота зрения – возможность различения двух соседних точек – максимальна для узких рецептивных полей центральной ямки. Слабые сигналы с периферии сетчатки также выделяются зрением, благодаря взаимодействию широких перекрывающихся рецептивных полей за счет пространственной суммации раздражителей.
В сетчатке есть клетки, которые наиболее эффективно реагируют на цвета-антагонисты: красный и зеленый, желтый и синий или зеленый и синий. Объясняется это явление тем, что из трех типов колбочек два всегда связаны с одной ганглиозной клеткой, часть колбочек имеет возбудительные синапсы, а часть – тормозные.
Сетчатка, по сложности организации рассматривается как часть мозга, расположенная на периферии. Здесь с фоторецепторами связано несколько слоев нейронов. Горизонтальные и биполярные клетки сетчатки не генерируют потенциалов действия. Они имеют градуальную гиперполяризацию и деполяризацию. Потенциалы действия генерируют ганглиозные клетки, аксоны которых образуют зрительный нерв, содержащий около 1 млн. волокон ганглиозных клеток сетчатки. Зрительные нервы обоих глаз перекрещиваются в области основания черепа, где одна половина волокон зрительного нерва переходит на противоположную сторону, а другая - вместе с перекрещенными аксонами второго зрительного нерва образуют зрительный тракт.
Нервные волокна зрительного тракта подходят к ядрам четырех структур мозга: верхние бугры четверохолмия и глазодвигательные ядра среднего мозга; латеральные коленчатые тела таламуса; супрахиазмальные ядра гипоталамуса.
В ядра верхних бугров четверохолмия и латерального коленчатого тела поступают ветви аксона одной и той же ганглиозной клетки сетчатки. Обе ветви сохраняют упорядоченную проекцию сетчатки. После переключения в переднем двухолмии сигналы поступают к ядру таламуса – подушке, а после переключения в латеральном коленчатом теле сигналы идут через зрительную радиацию и проецируются к клеткам первичной зрительной коры (поле 17 или стриарная кора). Проекция зоны максимальной остроты зрения сетчатки в 35 раз больше проекции участка такого же размера на периферии сетчатки. Клетки поля 17 (стриарной коры) связаны с вторичными зрительными зонами (поля 18 и 19, престриарная кора). От этих зон пути возвращаются к подушке таламуса, куда поступает информация от верхних бугров четверохолмия.
Клетки латерального коленчатого тела имеют простые концентрические рецептивные поля. Волокна от обоих глаз распределены топографически правильно и послойно, обеспечивая бинокулярное зрение. Небольшая часть клеток латерального коленчатого тела активируется от обоих зрительных нервов.
Нейроны зрительной коры имеют почти прямоугольные зрительные поля и функционально разделены на простые и сложные. Простые нейроны реагируют на световое пятно и имеют рецептивное поле, которое состоит из возбудительной и тормозной зоны. Сложные нейроны служат «детекторами» угла, наклона или движения линий в поле зрения. Наконец, кора имеет бинокулярную конвергенцию, так как в одной точке представлены симметричные поля зрения – справа и слева.
Распознаванию зрительных образов способствуют движения глаз. Движения глаз в одном направлении называются содружественными. При переводе взора с ближней точки ясного видения на дальнюю осуществляется дивергентные движения. При наклоне головы в сторону наблюдаются небольшие вращательные движения глаз.
При рассматривании предмета глаза двигаются от одной точки фиксации к другой быстрыми скачками – саккадами. Длительность саккад – от 10 до 80 мс, время периодов фиксации – 150 – 300 мс. При слежении за движущимися объектами возникают медленные (следящие) движения глаз.
Движения глаз управляются подкорковыми центрами ретикулярной формации, верхних бугров четверохолмия и претектальной области. Все эти центры координируются сигналами из зрительной, теменной и лобной коры, которые программируют движения тела и оценивают его положения в пространстве. Тонкую регуляцию глазодвигательных функций обеспечивает мозжечок, сравнивающий тонический и фазный компоненты движения при ориентации в пространстве.
При слежении за движущимся объектом возникает оптический нистагм, состоящий из чередования саккад и медленных следящих движений. При неподвижном глазном яблоке восприятие изображения исчезает в связи с разложением пигмента и адаптацией фоторецепторов.
Координированные движения глаз обеспечивают объединение информации, идущей от обоих глаз в центры мозга. Особое значение для координации движений играют нейроны переднего двухолмия. Во-первых, нейроны воспринимают сигналы, поступающие от одних и тех же участков полей зрения. Во-вторых, нейроны, на которых конвергирует импульсация от левого и правого глаз, являются пусковым механизмом для глазодвигательных нейронов. В коре имеются колонки, осуществляющие зрительное восприятие и сенсомоторную интеграцию, поэтому на высшем уровне зрительной системы параллельно функционируют две системы анализа. Одна определяет место предмета в пространстве, другая описывает его признаки. Конечные результаты параллельных процессов интегрируются и возникает законченный зрительный образ внешнего предметного мира.
Смирнов В.М., Будылина С.М. Физиология сенсорных систем и высшая нервная деятельность: Учеб. пособие для студ. высш. учеб. заведений. – М.: Академия, 2003. – с. 35 – 67
Зрительный анализатор
Зрительный анализатор представляет собой совокупность структур, воспринимающих световую энергию в виде электромагнитного излучения с длиной волны 400 – 700 нм и дискретных частиц фотонов, или квантов, и формирующих зрительные ощущения. С помощью глаза воспринимается 80 – 90 % всей информации об окружающем мире.

Рисунок: Орган зрения
Благодаря деятельности зрительного анализатора различают освещенность предметов, их цвет, форму, величину, направление передвижения, расстояние, на которое они удалены от глаза и друг от друга. Все это позволяет оценивать пространство, ориентироваться в окружающем мире, выполнять различные виды целенаправленной деятельности.
Наряду с понятием зрительного анализатора существует понятие органа зрения.
Орган зрения — это глаз, включающий три различных в функциональном отношении элемента:
1) глазное яблоко, в котором расположены световоспринимающий, светопреломляющий и светорегулирующий аппараты;
2) защитные приспособления, т. е. наружные оболочки глаза (склера и роговица), слезный аппарат, веки, ресницы, брови;
3) двигательный аппарат, представленный тремя парами глазных мышц (наружная и внутренняя прямые, верхняя и нижняя прямые, верхняя и нижняя косые), которые иннервируются III (глазодвигательный нерв), IV (блоковый нерв) и VI (отводящий нерв) парами черепных нервов.
Структурно-функциональная характеристика
Рецепторный (периферический) отдел зрительного анализатора (фоторецепторы) подразделяется на палочковые и колбочковые нейросенсорные клетки, наружные сегменты которых имеют соответственно палочковидную («палочки») и колбочковидную («колбочки») формы. У человека насчитывается 6 – 7 млн колбочек и 110 – 125 млн палочек.
Место выхода зрительного нерва из сетчатки не содержит фоторецепторов и называется слепым пятном. Латерально от слепого пятна в области центральной ямки лежит участок наилучшего видения — желтое пятно, содержащее преимущественно колбочки. К периферии сетчатки число колбочек уменьшается, а число палочек возрастает, и периферия сетчатки содержит одни лишь палочки.
Различия функций колбочек и палочек лежит в основе феномена двойственности зрения. Палочки являются рецепторами, воспринимающими световые лучи в условиях слабой освещенности, т.е. бесцветное, или ахроматическое, зрение. Колбочки же функционируют в условиях яркой освещенности и характеризуются разной чувствительностью к спектральным свойствам света (цветное или хроматическое зрение). Фоторецепторы обладают очень высокой чувствительностью, что обусловлено особенностью строения рецепторов и физико-химических процессов, лежащих в основе восприятия энергии светового стимула. Полагают, что фоторецепторы возбуждаются при действии на них 1 – 2 квантов света.
Палочки и колбочки состоят из двух сегментов – наружного и внутреннего, которые соединяются между собой посредством узкой реснички. Палочки и колбочки ориентированы в сетчатке радиально, а молекулы светочувствительных белков расположены в наружных сегментах таким образом, что около 90 % их светочувствительных групп лежат в плоскости дисков, входящих в состав наружных сегментов. Свет оказывает наибольшее возбуждающее действие в том случае, если направление луча совпадает с длинной осью палочки или колбочки, при этом он направлен перпендикулярно дискам их наружных сегментов.
Фотохимические процессы в сетчатке глаза. В рецепторных клетках сетчатки находятся светочувствительные пигменты (сложные белковые вещества) – хромопротеиды, которые обесцвечиваются на свету. В палочках на мембране наружных сегментов содержится родопсин, в колбочках – йодопсин и другие пигменты.
Родопсин и йодопсин состоят из ретиналя (альдегида витамина А1) и гликопротеида (опсина). Имея сходство в фотохимических процессах, они различаются тем, что максимум поглощения находится в различных областях спектра. Палочки, содержащие родопсин, имеют максимум поглощения в области 500 нм. Среди колбочек различают три типа, которые отличаются максимумами в спектрах поглощения: одни имеют максимум в синей части спектра (430 – 470 нм), другие в зеленой (500 – 530), третьи – в красной (620 – 760 нм) части, что обусловлено наличием трех типов зрительных пигментов. Красный кол бочковый пигмент получил название «йодопсин». Ретиналь может находиться в различных пространственных конфигурациях (изомерных формах), но только одна из них – 11-ЦИС-изомер ретиналя выступает в качестве хромофорной группы всех известных зрительных пигментов. Источником ретиналя в организме служат каротиноиды.
Фотохимические процессы в сетчатке протекают весьма экономно. Даже при действии яркого света расщепляется только небольшая часть имеющегося в палочках родопсина (около 0,006 %).
В темноте происходит ресинтез пигментов, протекающий с поглощением энергии. Восстановление йодопсина протекает в 530 раз быстрее, чем родопсина. Если в организме снижается содержание витамина А, то процессы ресинтеза родопсина ослабевают, что приводит к нарушению сумеречного зрения, так называемой куриной слепоте. При постоянном и равномерном освещении устанавливается равновесие между скоростью распада и ресинтеза пигментов. Когда количество света, падающего на сетчатку, уменьшается, это динамическое равновесие нарушается и сдвигается в сторону более высоких концентраций пигмента. Этот фотохимический феномен лежит в основе темновой адаптации.
Особое значение в фотохимических процессах имеет пигментный слой сетчатки, который образован эпителием, содержащим фусцин. Этот пигмент поглощает свет, препятствуя отражению и рассеиванию его, что обусловливает четкость зрительного восприятия. Отростки пигментных клеток окружают светочувствительные членики палочек и колбочек, принимая участие в обмене веществ фоторецепторов и в синтезе зрительных пигментов.
Вследствие фотохимических процессов в фоторецепторах глаза при действии света возникает рецепторный потенциал, который представляет собой гиперполяризацию мембраны рецептора. Это отличительная черта зрительных рецепторов, активация других рецепторов выражается в виде деполяризации их мембраны. Амплитуда зрительного рецепторного потенциала увеличивается при увеличении интенсивности светового стимула. Так, при действии красного цвета, длина волны которого составляет 620 – 760 нм, рецепторный потенциал более выражен в фоторецепторах центральной части сетчатки, а синего (430 – 470 нм) – в периферической.
Синаптические окончания фоторецепторов конвергируют на биполярные нейроны сетчатки. При этом фоторецепторы центральной ямки связаны только с одним биполяром.
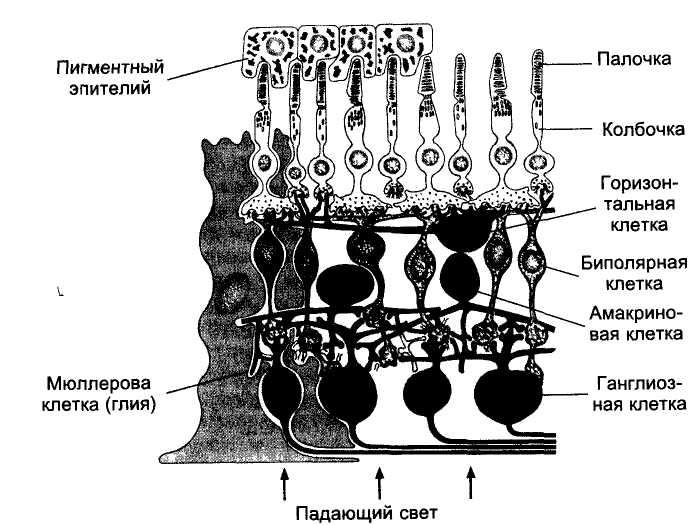
Рисунок: Схема строения сетчатки
Проводниковый отдел. Первый нейрон проводникового отдела зрительного анализатора представлен биполярными клетками сетчатки.
Считают, что в биполярных клетках возникают потенциалы действия подобно рецепторным и горизонтальным НС. В одних биполярах на включение и выключение света возникает медленная длительная деполяризация, а в других — на включение — гиперполяризация, на выключение — деполяризация.
Аксоны биполярных клеток в свою очередь конвергируют на ган-глиозные клетки (второй нейрон). В результате на каждую ганглиозную клетку могут конвергировать около 140 палочек и 6 колбочек, при этом чем ближе к желтому пятну, тем меньше фоторецепторов конвергирует на одну клетку. В области желтого пятна конвергенция почти не осуществляется и количество колбочек почти равно количеству биполярных и ганглиозных клеток. Именно это объясняет высокую остроту зрения в центральных отделах сетчатки.
Периферия сетчатки отличается большой чувствительностью к слабому свету. Это обусловлено, по-видимому, тем, что до 600 палочек конвергируют здесь через биполярные клетки на одну и ту же ганглиозную клетку. В результате сигналы от множества палочек суммируются и вызывают более интенсивную стимуляцию этих клеток.
[…]
Проводниковый отдел, начинающийся в сетчатке (первый нейрон — биполярный, второй нейрон — ганглиозные клетки), анатомически представлен далее зрительными нервами и после частичного перекреста их волокон — зрительными трактами. В каждом зрительном тракте содержатся нервные волокна, идущие от внутренней (носовой) поверхности сетчатки глаза одноименной стороны и от наружной половины сетчатки другого глаза. Волокна зрительного тракта направляются к зрительному бугру (собственно таламур), к метаталамусу (наружные коленчатые тела) и к ядрам подушки. Здесь расположены третий нейрон зрительного анализатора. От них зрительные нервные волокна направляются в кору полушарий большого мозга.
В наружных (или латеральных) коленчатых телах, куда приходят волокна из сетчатки, есть рецептивные поля, которые также имеют округлую форму, но меньше по размеру, чем в сетчатке. Ответы нейронов здесь носят фазический характер, но более выражены, чем в сетчатке. На уровне наружных коленчатых тел происходит процесс взаимодействия афферентных сигналов, идущих от сетчатки глаза, с эфферентными из области коркового отдела зрительного анализатора. С участием ретикулярной формации здесь происходит взаимодействие со слуховой и другими сенсорными системами, что обеспечивает процессы избирательного зрительного внимания путем выделения наиболее существенных компонентов сенсорного сигнала.
Центральный, или корковый отдел зрительного анализатора расположен в затылочной доле (поля 17, 18, 19 по Бродману) или VI, V2, V3 (согласно принятой номенклатуре). Считают, что первичная проекционная область (поле 17) осуществляет специализированную, но более сложную, чем в сетчатке и в наружных коленчатых телах, переработку информации. Рецептивные поля нейронов зрительной коры небольших размеров имеют вытянутые, почти прямоугольные, а не округлые формы. Наряду с этим имеются сложные и сверхсложные рецептивные поля детекторного типа. Эта особенность позволяет выделять из цельного изображения лишь отдельные части линий с различным расположением и ориентацией, при этом проявляется способность избирательно реагировать на эти фрагменты.
В каждом участке коры сконцентрированы нейроны, которые образуют колонку, проходящую по глубине через все слои вертикально, при этом происходит функциональное объединение нейронов, выполняющих сходную функцию. Разные свойства зрительных объектов (цвет, форма, движение) обрабатываются в разных частях зрительной коры большого мозга параллельно.
[…]
Механизмы, обеспечивающие ясное видение в различных условиях
При рассмотрении объектов, находящихся на разном удалении от наблюдателя, ясному видению способствуют следующие процессы.
1. Конвергенционные и дивергенционные движения глаз, благодаря которым осуществляется сведение или разведение зрительных осей. Если оба глаза двигаются в одном направлении, такие движения называются содружественными.
2. Реакция зрачка, которая происходит синхронно с движением глаз. Так, при конвергенции зрительных осей, когда рассматриваются близко расположенные предметы, происходит сужение зрачка, т. е. конвергентная реакция зрачков. Эта реакция способствует уменьшению искажения изображения, вызываемого сферической аберрацией. Сферическая аберрация обусловлена тем, что преломляющие среды глаза имеют неодинаковое фокусное расстояние в разных участках. Центральная часть, через которую проходит оптическая ось, имеет большее фокусное расстояние, чем периферическая часть. Поэтому изображение на сетчатке получается нерезким. Чем меньше диаметр зрачка, тем меньше искажения, вызываемые сферической аберрацией. Конвергентные сужения зрачка включают в действие аппарат аккомодации, обусловливающий увеличение преломляющей силы хрусталика.
Зрачок является также аппаратом устранения хроматической аберрации, которая обусловлена тем, что оптический аппарат глаза, как и простые линзы, преломляет свет с короткой волной сильнее, чем с длинной волной. Исходя из этого, для более точкой фокусировки предмета красного цвета требуется большая степень аккомодации, чем для синего. Именно поэтому синие предметы кажутся более удаленными, чем красные, будучи расположенными на одном и том же расстоянии.
3. Аккомодация является главным механизмом, обеспечивающим ясное видение разноудаленных предметов, и сводится к фокусированию изображения от далеко или близко расположенных предметов на сетчатке. Основной механизм аккомодации заключается в непроизвольном изменении кривизны хрусталика глаза.
Благодаря изменению кривизны хрусталика, особенно передней поверхности, его преломляющая сила может меняться в пределах 10 — 14 диоптрий. Хрусталик заключен в капсулу, которая по краям (вдоль экватора хрусталика) переходит в фиксирующую хрусталик связку (циннова связка), в свою очередь соединенную с волокнами ресничной (цилиарной) мышцы. При сокращении цилиарной мышцы натяжение цинновых связок уменьшается, а хрусталик вследствие своей эластичности становится более выпуклым. Преломляющая сила глаза увеличивается, и глаз настраивается на видение близко расположенных предметов. Когда человек смотрит вдаль, циннова связка находится в натянутом состоянии, что приводит к растягиванию сумки хрусталика и его утолщению. Иннервация цилиарной мышцы осуществляется симпатическими и парасимпатическими нервами. Импульсация, поступающая по парасимпатическим волокнам глазодвигательного нерва, вызывает сокращение мышцы. Симпатические волокна, отходящие от верхнего шейного узла, вызывают ее расслабление. Изменение степени сокращения и расслабления цилиарной мышцы связано с возбуждением сетчатки и находится под влиянием коры головного мозга. Преломляющая сила глаза выражается в диоптриях (Д). Одна диоптрия соответствует преломляющей силе линзы, главное фокусное расстояние которой в воздухе равно 1 м. Если главное фокусное расстояние линзы равно, например, 0,5 или 2 м, то ее преломляющая сила составляет, соответственно, 2Д или 0,5Д. Преломляющая сила глаза без явления аккомодации равна 58 —60 Д и называется рефракцией глаза.
При нормальной рефракции глаза лучи от далеко расположенных предметов после прохождения через светопреломляющую систему глаза собираются в фокусе на сетчатке в центральной ямке. Нормальная рефракция глаза носит название эмметропии, а такой глаз называют эмметропическим. Наряду с нормальной рефракцией наблюдаются ее аномалии.
Миопия (близорукость) — это такой вид нарушения рефракции, при котором лучи от предмета после прохождения через светопреломляющий аппарат фокусируются не на сетчатке, а впереди нее. Это может зависеть от большой преломляющей силы глаза или от большой длины глазного яблока. Близкие предметы близорукий видит без аккомодации, отдаленные предметы видит неясными, расплывчатыми. Для коррекции применяются очки с рассеивающими двояковогнутыми линзами.
Гиперметропия (дальнозоркость) — вид нарушения рефракции, при котором лучи от далеко расположенных предметов в силу слабой преломляющей способности глаза или при малой длине глазного яблока фокусируются за сетчаткой. Даже удаленные предметы дальнозоркий глаз видит с напряжением аккомодации, вследствие чего развивается гипертрофия аккомодационных мышц. Для коррекции применяют двояковыпуклые линзы.
Астигматизм — вид нарушения рефракции, при котором лучи не могут сходиться в одной точке, в фокусе (от греч. stigme — точка), обусловлен различной кривизной роговицы и хрусталика в различных меридианах (плоскостях). При астигматизме предметы кажутся сплющенными или вытянутыми, его коррекцию осуществляют сфероцилиндрическими линзами.
Следует отметить, что к светопреломляющей системе глаза относятся также: роговица, влага передней камеры глаза, хрусталик и стекловидное тело. Однако их преломляющая сила, в отличие от хрусталика, не регулируется и в аккомодации участия не принимает. После прохождения лучей через преломляющую систему глаза на сетчатке получается действительное, уменьшенное и перевернутое изображение. Но в процессе индивидуального развития сопоставление ощущений зрительного анализатора с ощущениями двигательного, кожного, вестибулярного и других анализаторов, как отмечалось выше, приводит к тому, что человек воспринимает внешний мир таким, какой он есть на самом деле.
Бинокулярное зрение (зрение двумя глазами) играет важную роль в восприятии разноудаленных предметов и определении расстояния до них, дает более выраженное ощущение глубины пространства по сравнению с монокулярным зрением, т. е. зрением одним глазом. При рассматривании предмета двумя глазами его изображение может попадать на симметричные (идентичные) точки сетчаток обоих глаз, возбуждения от которых объединяются в корковом конце анализатора в единое целое, давая при этом одно изображение. Если изображение предмета попадает на неидентичные (диспаратные) участки сетчатки, то возникает раздвоение изображения. Процесс зрительного анализа пространства зависит не только от наличия бинокулярного зрения, существенную роль в этом играют условно-рефлекторные взаимодействия, складывающиеся между зрительным и двигательным анализаторами. Определенное значение имеют конвергенционные движения глаз и процесс аккомодации, которые управляются по принципу обратный связей. Восприятие пространства в целом связано с определением пространственных отношений видимых предметов — их величины, формы, отношения друг к другу, что обеспечивается взаимодействием различных отделов анализатора; значительную роль при этом играет приобретенный опыт.
При движении объектов ясному видению способствуют следующие факторы: 1) произвольные движения глаз вверх, вниз, влево или вправо со скоростью движения объекта, что осуществляется благодаря содружественной деятельности глазодвигательных мышц; 2) при появлении объекта в новом участке поля зрения срабатывает фиксационный рефлекс — быстрое непроизвольное движение глаз, обеспечивающее совмещение изображения предмета на сетчатке с центральной ямкой. При слежении за движущимся объектом происходит медленное движение глаз — следящее движение.
При рассматривании неподвижного предмета для обеспечения ясного видения глаз совершает три типа мелких непроизвольных Движений: тремор — дрожание глаза с небольшой амплитудой и частотой, дрейф — медленное смещение глаза на довольно значительное расстояние и скачки (флики) — быстрые движения глаз. Также существуют саккадические движения (саккады) — содружественные движения обоих глаз, совершаемые с большой скоростью. Наблюдаются саккады при чтении, просматривании картин, когда обследуемые точки зрительного пространства находятся на одном удалении 6т наблюдателя и других объектов. Если заблокировать эти движения глаз, то окружающий нас мир вследствие адаптации рецепторов сетчатки станет трудно различимым, каким он является у лягушки. Глаза лягушки неподвижны, поэтому она хорошо различает только движущиеся предметы, например бабочек. Именно поэтому лягушка приближается к змее, которая постоянно выбрасывает наружу свой язык. Находящуюся в состоянии неподвижности змею лягушка не различает, а ее движущийся язык принимает за летающую бабочку.
В условиях изменения освещенности ясное видение обеспечивают зрачковый рефлекс, темновая и световая адаптация.
Зрачок регулирует интенсивность светового потока, действующего на сетчатку, путем изменения своего диаметра. Ширина зрачка может колебаться от 1,5 до 8,0 мм. Сужение зрачка (миоз) происходит при увеличении освещенности, а также при рассматривании близко расположенного предмета и во сне. Расширение зрачка (мидриаз) происходит при уменьшении освещенности, а также при возбуждении рецепторов, любых афферентных нервов, при эмоциональных реакциях напряжения, связанных с повышением тонуса симпатического отдела нервной системы (боль, гнев, страх, радость и т.д.), при психических возбуждениях (психозы, истерии и т.д.), при удушье, наркозе. Зрачковый рефлекс при изменении освещенности хотя и улучшает зрительное восприятие (в темноте расширяется, что увеличивает световой поток, падающий на сетчатку, на свету сужается), однако главным механизмом все же является темновая и световая адаптация.
Темновая адаптация выражается в повышении чувствительности зрительного анализатора (сенситизация), световая адаптация — в снижении чувствительности глаза к свету. Основу механизмов световой и темновой адаптации составляют протекающие в колбочках и палочках фотохимические процессы, которые обеспечивают расщепление (на свету) и ресинтез (в темноте) фоточувствительных пигментов, а также процессы функциональной мобильности: включение и выключение из деятельности рецепторных элементов сетчатки. Кроме того, адаптацию определяют некоторые нейронные механизмы и, прежде всего, процессы, происходящие в нервных элементах сетчатки, в частности способы подключения фоторецепторов к ганглиозным клеткам с участием горизонтальных и биполярных клеток. Так, в темноте возрастает число рецепторов, подключенных к одной биполярной клетке, и большее их число конвергирует на ганглиозную клетку. При этом расширяется рецептивное поле каждой биполярной и, естественно, ганглиозной клеток, что улучшает зрительное восприятие. Включение же горизонтальных клеток регулируется ЦНС.
Снижение тонуса симпатической нервной системы (десимпатизация глаза) уменьшает скорость темновой адаптации, а введение адреналина оказывает противоположный эффект. Раздражение ретикулярной формации ствола мозга повышает частоту импульсов в волокнах зрительных нервов. Влияние ЦНС на адаптивные процессы в сетчатке подтверждается также тем, что чувствительность неосвещенного глаза к свету изменяется при освещении другого глаза и при действии звуковых, обонятельных или вкусовых раздражителей.
Цветовая адаптация. Наиболее быстрая и резкая адаптация (снижение чувствительности) происходит при действии сине-фиолетового раздражителя. Красный раздражитель занимает среднее положение.
Зрительное восприятие крупных объектов и их деталей обеспечивается за счет центрального и периферического зрения — изменений угла зрения. Наиболее тонкая оценка мелких деталей предмета обеспечивается в том случае, если изображение падает на желтое пятно, которое локализуется в центральной Ямке сетчатки глаза, так как в этом случае имеет место наибольшая острота зрения. Это объясняется тем, что в области желтого пятна располагаются только колбочки, их размеры наименьшие, и каждая колбочка контактирует с малым числом нейронов, что повышает
остроту зрения. Острота зрения определяется наименьшим углом зрения, под которым глаз еще способен видеть отдельно две точки. Нормальный глаз способен различать две светящиеся точки под углом зрения в 1’. Острота зрения такого глаза принимается за единицу. Острота зрения зависит от оптических свойств глаза, структурных особенностей сетчатки и работы нейрональных механизмов проводникового и центрального отделов зрительного анализатора. Определение остроты зрения осуществляется с помощью буквенных или различного вида фигурных стандартных таблиц. Крупные объекты в целом и окружающее пространство воспринимаются в основном за счет периферического зрения, обеспечивающего большое поле зрения.
Поле зрения — пространство, которое можно видеть фиксированным глазом. Различают отдельно поле зрения левого и правого глаз, а также общее поле зрения для двух глаз. Величина поля зрения у людей зависит от глубины положения глазного яблока и формы надбровных дуг и носа. Ахроматическое поле зрения больше хроматического в силу того, что на периферии сетчатки нет рецепторов, воспринимающих цвет (колбочек). В свою очередь, цветное поле зрения неодинаково для различных цветов. Самое узкое поле зрения для зеленого, желтого, больше для красного, еще больше для синего цветов. Величина поля зрения изменяется в зависимости от освещенности. Ахроматическое поле зрения в сумерках увеличивается, на свету уменьшается. Хроматическое поле зрения, наоборот, на свету увеличивается, в сумерках уменьшается. Это зависит от процессов мобилизации и демобилизации фоторецепторов (функциональной мобильности). При сумеречном зрении увеличение количества функционирующих палочек, т.е. их мобилизация, приводит к увеличению ахроматического поля зрения, в то же самое время уменьшение количества функционирующих колбочек (их демобилизация) ведет к уменьшению хроматического поля зрения.
Зрительный анализатор имеет также механизм для различения длины световой волны — цветовое зрение.
Цветовое зрение, зрительные контрасты и последовательные образы
Цветовое зрение — способность зрительного анализатора реагировать на изменения длины световой волны с формированием ощущения цвета. Определенной длине волны электромагнитного излучения соответствует ощущение определенного цвета. Так, ощущение красного цвета соответствует действию света с длиной волны в 620 — 760 нм, а фиолетового — 390 — 450 нм, остальные цвета спектра имеют промежуточные параметры. Смешение всех цветов дает ощущение белого цвета. В результате смешения трех основных цветов спектра — красного, зеленого, сине-фиолетового — в разном соотношении можно получить также восприятие любых других цветов. Ощущение цветов связано с освещенностью. По мере ее уменьшения сначала перестают различаться красные цвета, позднее всех — синие. Восприятие цвета обусловлено в основном процессами, происходящими в фоторецепторах. Наибольшим признанием пользуется трехкомпонентная теория цветоощущения Ломоносова— Юнга—Гельмгольца—Лазарева, согласно которой в сетчатке глаза имеются три вида фоторецепторов — колбочек, раздельно воспринимающих красный, зеленый и сине-фиолетовые цвета. Комбинации возбуждения различных колбочек приводят к ощущению различных цветов и оттенков. Равномерное возбуждение трех видов колбочек дает ощущение белого цвета. Три типа цветочувствительных колбочек были названы модуляторами, колбочки, которые возбуждались при изменении яркости света (четвертый тип), были названы доминаторами. Впоследствии методом микроспектрофотометрии удалось установить, что даже одиночная колбочка может поглощать лучи различной длины волны. Обусловлено это наличием в каждой колбочке различных пигментов, чувствительных к волнам света различной длины.
Несмотря на убедительные аргументы трехкомпонентной теории в физиологии цветового зрения описаны факты, которые не находят объяснения с этих позиций. Это дало возможность выдвинуть теорию противоположных, или контрастных, цветов, т. е. создать так называемую оппонентную теорию цветного зрения Эвальда Геринга.
Согласно этой теории, в глазу и/или в мозге существуют три оппонентных процесса: один — для ощущения красного и зеленого, второй — для ощущения желтого и синего, третий — качественно отличный от двух первых процессов — для черного и белого. Эта теория применима для объяснения передачи информации о цвете в последующих отделах зрительной системы: ганглиозных клетках сетчатки, наружных коленчатых телах, корковых центрах зрения, где функционируют цветооппонентные РП с их центром и периферией.
Таким образом, на основании полученных данных можно полагать, что процессы в колбочках более соответствуют трехкомпонентной теории цветоощущения, тогда как для нейронных сетей сетчатки и вышележащих зрительных центров подходит теория контрастных цветов Геринга.
В восприятии цвета определенную роль играют и процессы, Протекающие в нейронах разных уровней зрительного анализатора (включая сетчатку), которые получили название цветооппонентных нейронов. При действии на глаз излучений одной части спектра они возбуждаются, а другой — тормозятся. Такие нейроны участвуют в кодировании информации о цвете.
Наблюдаются аномалии цветового зрения, которые могут проявляться в виде частичной или полной цветовой слепоты. Людей, вообще не различающих цвета, называют ахроматами. Частичная цветовая слепота имеет место у 8 – 10% мужчин и 0,5% женщин. Полагают, что цветослепота связана с отсутствием у мужчин определенных генов в половой непарной Х-хромосоме. Различаются три вида частичной цветослепоты: протанопия (дальтонизм) — слепота в основном на красный цвет. Этот вид цветослепоты впервые был описан в 1794 году физиком Дж. Дальтоном, у которого наблюдался этот вид аномалии. Людей с таким видом аномалии называют «краснослепыми»; дейтеранопия — понижение восприятия зеленого цвета. Таких людей называют «зеленослепыми»; тританопия — редко встречающаяся аномалия. При этом люди не воспринимают синий и фиолетовый цвета, их называют «фиолетовослепыми».
С точки зрения трехкомпонентной теории цветового зрения каждый из видов аномалии является результатом отсутствия одного из трех колбочковых цветовоспринимающих субстратов. Для диагностики расстройства цветоощущения пользуются цветными таблицами Е.Б. Рабкина, а также специальными приборами, получившими название аномалоскопов. Выявление различных аномалий цветового зрения имеет большое значение при определении профессиональной пригодности человека для различных видов работ (шофера, летчика, художника и др.).
Возможность оценки длины световой волны, проявляющаяся в способности к цветоощущению, играет существенную роль в жизни человека, оказывая влияние на эмоциональную сферу и деятельность различных систем организма. Красный цвет вызывает ощущение тепла, действует возбуждающе на психику, усиливает эмоции, но быстро утомляет, приводит к напряжению мышц, повышению артериального давления, учащению дыхания. Оранжевый цвет вызывает чувство веселья и благополучия, способствует пищеварению. Желтый цвет создает хорошее, приподнятое настроение, стимулирует зрение и нервную систему. Это самый «веселый» цвет. Зеленый цвет действует освежающе и успокаивающе, полезен при бессоннице, переутомлении, понижает артериальное давление, общий тонус организма и является самым благоприятным для человека. Голубой цвет вызывает ощущение прохлады и действует на нервную систему успокаивающе, причем сильнее зеленого (особенно благоприятен голубой цвет для людей с повышенной нервной возбудимостью), больше, чем при зеленом цвете, понижает артериальное давление и тонус мышц. Фиолетовый цвет не столько успокаивает, сколько расслабляет психику. Создается впечатление, что человеческая психика, следуя вдоль спектра от красного к фиолетовому, проходит всю гамму эмоций. На этом основано использование теста Люшера для определения эмоционального состояния организма.
Зрительные контрасты и последовательные образы. Зрительные ощущения могут продолжаться и после того, как прекратилось раздражение. Такое явление получило название последовательных образов. Зрительные контрасты — это измененное восприятие раздражителя в зависимости от окружающего светового или цветового фона. Существуют понятий светового и цветового зрительных контрастов. Явление контраста может проявляться в преувеличении действительной разницы между двумя одновременными или последовательными ощущениями, поэтому различают одновременные и последовательные контрасты. Серая полоска на белом фоне кажется темнее такой же полоски, расположенной на темном фоне. Это пример одновременного светового контраста. Если рассматривать серый цвет на красном фоне, то он кажется зеленоватым, а если рассматривать серый цвет на синем фоне, то он приобретает желтый оттенок. Это явление одновременного цветового контраста. Последовательный цветовой контраст заключается в изменении цветового ощущения при переводе взгляда на белый фон. Так, если долго смотреть на окрашенную в красный цвет поверхность, а затем перевести взор на белую, то она приобретает зеленоватый оттенок. Причиной зрительного контраста являются процессы, которые осуществляются в фоторецепторном и нейрональном аппаратах сетчатки. Основу составляет взаимное торможение клеток, относящихся к разным рецептивным полям сетчатки и их проекциям в корковом отделе анализаторов.
Психофизиология. Под ред. Ю.И. Александрова. – СПб.: Питер, 2003
Зрительная система
Зрение — одно из важнейших чувств человека. Оно эволюционно приспособлено к восприятию узкой части диапазона электромагнитных излучений (видимый свет). Зрительная система дает мозгу более 90 % сенсорной информации. Зрение — это многозвеньевой процесс, начинающийся с проекции изображения на сетчатую оболочку глаза. Затем происходит возбуждение фоторецепторов, передача и преобразование зрительной информации в нейронных слоях зрительной системы, а заканчивается зрительное восприятие принятием высшими корковыми отделами зрительной системы решения о зрительном образе.
Строение и функции оптического аппарата глаза
Глазное яблоко имеет шарообразную форму, что облегчает его повороты для наведения на рассматриваемый объект и обеспечивает хорошую фокусировку изображения на всей светочувствительной оболочке глаза — сетчатке. На пути к сетчатке лучи света проходят через несколько прозрачных сред — роговицу, хрусталик и стекловидное тело. Определенная кривизна и показатель преломления роговицы и в меньшей мере хрусталика определяют преломление световых лучей внутри глаза. На сетчатке получается изображение, резко уменьшенное и перевернутое вверх ногами и справа налево. Преломляющую силу любой оптической системы выражают в диоптриях. Одна диоптрия равна преломляющей силе линзы с фокусным расстоянием 100 см […]
Аккомодация
Аккомодацией называют приспособление глаза к ясному видению объектов, расположенных на разном расстоянии (подобно фокусировке в фотографии). Для ясного видения объекта необходимо, чтобы его изображение было сфокусировано на сетчатке. Главную роль в аккомодации играет изменение кривизны хрусталика, т. е. его преломляющей способности. При рассматривании близких предметов хрусталик становится более выпуклым. Механизмом аккомодации является сокращение мышц, изменяющих выпуклость хрусталика.
[…]
Зрачок и зрачковый рефлекс
Зрачок — это отверстие в центре радужной оболочки, через которое свет проходит в глаз. Он повышает четкость изображения на сетчатке, увеличивая глубину резкости глаза и устраняя сферическую аберрацию. Расширившийся при затемнении зрачок на свету быстро сужается («зрачковый рефлекс»), что регулирует поток света, попадающий в глаз. Так, на ярком свету зрачок имеет диаметр 1,8 мм, при средней дневной освещенности он расширяется до 2,4 мм, а в темноте — до 7,5 мм. Это ухудшает качество изображения на сетчатке, но увеличивает абсолютную чувствительность зрения. Реакция зрачка на изменение освещенности имеет адаптивный характер, так как стабилизирует освещенность сетчатки в небольшом диапазоне. У здоровых людей зрачки обоих глаз имеют одинаковый диаметр. При освещении одного глаза зрачок другой тоже суживается; подобная реакция называется содружественной.
Структура и функции сетчатки
Сетчатка — это внутренняя светочувствительная оболочка глаза. Она имеет сложную многослойную структуру. Здесь расположены два вида фоторецепторов (палочки и колбочки) и несколько видов нервных клеток. Возбуждение фоторецепторов активирует первую нервную клетку сетчатки — биполярный нейрон. Возбуждение биполярных нейронов активирует ганглиозные клетки сетчатки, передающие свои импульсы в подкорковые зрительные центры. В процессах передачи и переработки информации в сетчатке участвуют также горизонтальные и амакриновые клетки. Все перечисленные нейроны сетчатки с их отростками образуют нервный аппарат глаза, который участвует в анализе и переработке зрит информации. Именно поэтому сетчатку называют частью мозга, вынесенной на периферию.
Структура и функции слоев сетчатки
Клетки пигментного эпителия образуют наружный, наиболее далекий от света, слой сетчатки. Они содержат меланосомы, придающие им черный цвет. Пигмент поглощает излишний свет, препятствуя его отражению и рассеиванию, что способствует четкости изображения на сетчатке. Пигментный эпителий играет решающую роль в регенерации зрительного пурпура фоторецепторов после его обесцвечивания, в постоянном обновлении наружных сегментов зрит клеток, в защите рецепторов от светового повреждения, а также в переносе к ним кислорода и питательных веществ.
Фоторецепторы. К слою пигментного эпителия изнутри примыкает слой зрит рецепторов: палочек и колбочек. В каждой сетчатке человека находится 6 – 7 млн. колбочек и 110 – 125 млн палочек. Они распределены в сетчатке неравномерно. Центральная ямка сетчатки — фовеа содержит только колбочки. По направлению к периферии сетчатки количество колбочек уменьшается, а количество палочек увеличивается, так что на дальней периферии имеются только палочки. Колбочки функционируют в условиях больших освещенностей, они обеспечивают дневное и цветовое зрение; более светочувствительные палочки ответственны за сумеречное зрение.
Цвет воспринимается лучше всего при действии света на центральную ямку сетчатки, в которой расположены почти исключительно колбочки. Здесь же и наибольшая острота зрения. По мере удаления от центра сетчатки восприятие цвета и пространственное разрешение постепенно уменьшается. Периферия сетчатки, на которой находятся исключительно палочки, не воспринимает цвета. Зато световая чувствительность колбочкового аппарата сетчатки во много раз меньше, чем у палочкового. Поэтому в сумерках из-за резкого понижения колбочкового зрения и преобладания периферического палочкового зрения мы не различаем цвет («ночью все кошки серы»).
Нейроны сетчатки
Фоторецепторы сетчатки синаитически связаны с биполярными нервными клетками. При действии света уменьшается выделение медиатора из фоторецептора, что гиперполяризует мембрану биполярной клетки. От нее нервный сигнал передается на ганглиозные клетки, аксоны которых являются волокнами зрительного нерва.
На 130 млн фоторецепторных клеток приходится только 1 млн 250 тыс. ганглиозных клеток сетчатки. Это значит, что импульсы от многих фоторецепторов сходятся (конвергируют) через биполярные нейроны к одной гаиглиозной клетке. Фоторецепторы, соединенные с одной ганглиозной клеткой, образуют ее рецептивное поле. Т.о., каждая ганглиозная клетка суммирует возбуждение, возникающее в большом количестве фоторецепторов. Это повышает световую чувствительность сетчатки, но ухудшает ее пространственное разрешение. Лишь в центре сетчатки (в районе центральной ямки) каждая колбочка соединена с одной биполярной клеткой, а та, в свою очередь, соединена с одной ганглиозной клеткой. Это обеспечивает высокое пространственное разрешение центра сетчатки, но резко уменьшает его световую чувствительность.
Взаимодействие соседних нейронов сетчатки обеспечивается горизонтальными и амакриновыми клетками, через отростки которых распространяются сигналы, меняющие синаптическую передачу между фоторецепторами и биполярами (горизонтальные клетки) и между биполярами и ганглиозными клетками (амакрины). Амакриновые клетки осуществляют боковое торможение между соседними ганглиозными клетками. В сетчатку приходят и центробежные, или эфферентные, нервные волокна, приносящие к ней сигналы из мозга. Эти импульсы регулируют проведение возбуждения между биполярными и ганглиозными клетками сетчатки.
Нервные пути и связи в зрительной системе
Из сетчатки зрит информация по волокнам зрит нерва устремляется в мозг. Нервы от двух глаз встречаются у основания мозга, где часть волокон переходит на противоположную сторону (зрительный перекрест, или хиазма). Это обеспечивает каждое полушарие мозга информацией от обоих глаз: в затылочную долю правого полушария поступают сигналы от правой половины каждой сетчатки, а в левое полушарие — от левой.
После хиазмы зрительные нервы называются оптическими трактами и основное количество их волокон приходит в подкорковый зрительный центр — наружное коленчатое тело (НКТ). Отсюда зрительные сигналы поступают в первичную проекционную область зрит коры (стриарная кора, или поле 17 по Бродману). Зрительная кора состоит из ряда полей, каждое из которых обеспечивает свои, специфические функции, получая как прямые, так и опосредованные сигналы от сетчатки и в общем сохраняя ее топологию, или ретинотопию (сигналы от соседних участков сетчатки попадают в соседние участки коры).
Импульсные разряды нейронов НКТ по их аксонам поступают в затылочную часть полушарий гм, в которой расположена первичная проекционная область зрит коры (стриарная кора). Здесь у приматов и человека происходит значительно более специализированная и сложная, чем в сетчатке и в НКТ, переработка информации. Нейроны зрит коры имеют не круглые, а вытянутые (по горизонтали, вертикали или по диагонали) рецептивные поля небольшого размера. Благодаря этому они способны выделять из изображения отдельные фрагменты линий с той или иной ориентацией и расположением и избирательно на них реагировать (детекторы ориентации). В каждом небольшом участке зрит коры по ее глубине сконцентрированы нейроны с одинаковой ориентацией и локализацией рецептивных полей в поле зрения. Они образуют ориентационную колонку нейронов, проходящую вертикально через все слои коры. Колонка — пример функционального объединения корковых нейронов, осуществляющих сходную функцию. Группа соседних ориентационных колонок, нейроны которых имеют перекрывающиеся рецептивные поля, но разные предпочитаемые ориентации, образует так называемую сверхколонку.

Рисунок: схема зрит путей от сетчатки глаза до первичной зрит коры:
ЛПЗ — левое поле зрения; ППЗ — правое поле зрения; тф — точка фиксации взора; лг — левый глаз; пг — правый глаз; зн — зрительный нерв; х — зрительный перекрест, или хиазма; от — оптический тракт; НКТ — наружное коленчатое тело; ЗК — зрительная кора; лп — левое полушарие; пп – правое полушарие
[…]
Цветовое зрение
Весь видимый нами спектр электромагнитных излучений заключен между коротковолновым (длина волны 400 нм) излучением, которое мы называем фиолетовым цветом, и длинноволновым излучением (длина волны 700 нм), называемым красным цветом. Остальные цвета видимого спектра (синий, зеленый, желтый и оранжевый) имеют промежуточные значения длины волны. Смешение лучей всех цветов дает белый цвет. Он может быть получен и при смешении двух так называемых парных дополнительных цветов: красного и синего, желтого и синего. Если произвести смешение трех основных цветов — красного, зеленого и синего, — то могут быть получены любые цвета.
Максимальным признанием пользуется трехкомпонентная теория Г. Гельмгольца, согласно которой цветовое восприятие обеспечивается тремя типами колбочек с различной цветовой чувствительностью. Одни из них чувствительны к красному цвету, другие — к зеленому, а третьи — к синему. Всякий цвет оказывает воздействие на все три цветоощущающих элемента, но в разной степени. Эта теория прямо подтверждена в опытах, в которых измеряли поглощение излучений с разной длиной волны в одиночных колбочках сетчатки человека.
Частичная цветовая слепота была описана в конце XVIII в. Д. Дальтоном, который сам страдал ею. Поэтому аномалию цветовосприятия обозначили термином «дальтонизм». Дальтонизм встречается у 8 % мужчин; его связывают с отсутствием определенных генов в определяющей пол непарной у мужчин Х-хромосоме. Для диагностики дальтонизма, важной при профессиональном отборе, используют полихроматические таблицы. Люди, страдающие им, не могут быть полноценными водителями транспорта, так как они могут не различать цвет огней светофоров и дорожных знаков. Существуют три разновидности частичной цветовой слепоты: протанопия, дейтеранопия и тританопия. Каждая из них характеризуется отсутствием восприятия одного из трех основных цветов. Люди, страдающие протано-пией («краснослепые»), не воспринимают красного цвета, сине-голубые лучи кажутся им бесцветными. Лица, страдающие дейтеранопией («зеленослепые»), не отличают зеленые цвета от темно-красных и голубых. При тританопии (редко встречающейся аномалии цветового зрения) не воспринимаются лучи синего и фиолетового цвета. Все перечисленные виды частичной цветовой слепоты хорошо объясняются трехкомпонентной теорией. Каждый из них является рез-том отсутствия одного из трех колбочковых цветовоспринимающих веществ.
Восприятие пространства
Остротой зрения называется максимальная способность различать отдельные детали объектов. Ее определяют по наименьшему расстоянию между двумя точками, которые различает глаз, т. е. видит отдельно, а не слитно. Нормальный глаз различает две точки, расстояние между которыми составляет 1 угловую минуту. Максимальную остроту зрения имеет центр сетчатки — желтое пятно. К периферии от него острота зрения намного меньше. Острота зрения измеряется при помощи специальных таблиц, которые состоят из нескольких рядов букв или незамкнутых окружностей различной величины. Острота зрения, определенная по таблице, выражается в относительных величинах, причем нормальная острота принимается за единицу. Встречаются люди, обладающие сверхостротой зрения.
[…]
Бинокулярное зрение — это зрение двумя глазами. При взгляде на какой-либо предмет у человека с нормальным зрением не возникает ощущения двух предметов, хотя и имеется два изображения на двух сетчатках. Изображение каждой точки этого предмета попадает на так называемые корреспондирующие, или соответственные, участки двух сетчаток, и в восприятии человека два изображения сливаются в одно. Если надавить слегка на один глаз сбоку, то начнет двоиться в глазах, потому что нарушилось соответствие сетчаток. Если же смотреть на близкий предмет, то изображение какой-либо более отдаленной точки попадает на неидентичные (диспаратные) точки двух сетчаток. Диспарация играет большую роль в оценке расстояния и, следовательно, в видении глубины пространства. […]
Оценка величины объекта. Величина знакомого предмета оценивается как функция величины его изображения на сетчатке и расстояния предмета от глаз. В случае, когда расстояние до незнакомого предмета оценить трудно, возможны грубые ошибки в определении его величины.
Оценка расстояния. Восприятие глубины пространства и оценка расстояния до объекта возможны при зрении как одним глазом (монокулярное зрение), так и двумя глазами (бинокулярное зрение). Во втором случае оценка расстояния гораздо точнее. Некоторое значение в оценке близких расстояний при монокулярном зрении имеет явление аккомодации. Для оценки расстояния имеет значение также то, что знакомый предмет кажется ближе.
Роль движения глаз для зрения. При рассматривании любых предметов глаза двигаются. Глазные движения осуществляют 6 мышц, прикрепленных к глазному яблоку. Движение двух глаз совершается одновременно и содружественно. Рассматривая близкие предметы, необходимо сводить (конвергенция), а рассматривая далекие предметы — разводить зрительные оси двух глаз (дивергенция). Кроме того, важная роль движений глаз для зрения определяется также тем, что для непрерывного получения мозгом зрит информации необходимо движение изображения на сетчатке. Импульсы в зрительном нерве возникают в момент включения и выключения светового изображения. При длящемся действии света на одни и те же фоторецепторы импульсация в волокнах зрительного нерва быстро прекращается и зрительное ощущение при неподвижных глазах и объектах исчезает через 1 – 2 с. Если на глаз поставить присоску с крохотным источником света, то человек видит его только в момент включения или выключения, так как этот раздражитель движется вместе с глазом и, следовательно, неподвижен по отношению к сетчатке. Чтобы преодолеть такое приспособление (адаптацию) к неподвижному изображению, глаз при рассматривании любого предмета производит не ощущаемые человеком непрерывные скачки (саккады). Вследствие каждого скачка изображение на сетчатке смещается с одних фоторецепторов на другие, вновь вызывая импульсацию ганглиозных клеток. Продолжительность каждого скачка равна сотым долям секунды, а амплитуда его не превышает 20 угловых градусов. Чем сложнее рассматриваемый объект, тем сложнее траектория движения глаз. Они как бы «прослеживают» контуры изображения, задерживаясь на наиболее информативных его участках. Кроме скачков, глаза непрерывно мелко дрожат и дрейфуют (медленно смещаются с точки фиксации взора). Эти движения также очень важны для зрительного восприятия.
Хомская Е.Д. Нейропсихология: 4-е издание. – СПб.: Питер, 2005. – 496 с.
Сенсорные и гностические зрительные расстройства. Зрительные агнозии
Общие принципы работы анализаторных систем
[…]
Анализаторные системы человека — сложные многоуровневые образования, направленные на анализ
сигналов определенной модальности.
Можно выделить несколько общих принципов строения всех анализаторных систем:
а) принцип параллельной многоканальной переработки информации, в соответствии с которым информация о разных параметрах сигнала одновременно передается по различным каналам анализаторной системы;
б) принцип анализа информации с помощью нейронов-детекторов, направленного на выделение как относительно элементарных, так и сложных, комплексных характеристик сигнала, что обеспечивается разными рецептивными полями;
в) принцип последовательного усложнения переработки информации от уровня к уровню, в соответствии с которым каждый из них осуществляет свои собственные анализаторные функции;
г) принцип топического («точка в точку») представительства периферических рецепторов в первичном поле анализаторной системы;
д) принцип целостной интегративной репрезентации сигнала в ЦНС во взаимосвязи с другими сигналами, что достигается благодаря существованию общей модели (схемы) сигналов данной модальности (по типу «сферической модели цветового зрения»).
[…] При нейропсихологическом изучении работы анализаторных систем следует различать два типа расстройств:
1) относительно элементарные сенсорные расстройства в виде нарушений различных видов ощущений (светоощущений, цветоощущений, ощущений высоты, громкости, длительности звука и др.);
2) более сложные гностические расстройства в виде нарушений разных видов восприятия (восприятия формы предмета, пространственных отношений, символов, звуков речи и т. д.).
Первый тип расстройств связан с поражением периферического и подкорковых уровней анализаторных систем, а также первичного коркового поля соответствующего анализатора.
Второй тип расстройств обусловлен прежде всего поражением вторичных корковых полей, хотя в мозговой организации гностической деятельности принимают участие и многие другие корковые и подкорковые структуры, в том числе префронтальные отделы коры больших полушарий.
Гностические расстройства, возникающие при корковых очагах поражения, носят название агнозий. В зависимости от пораженного анализатора различают зрительные, слуховые и тактильные агнозии.
Важно отметить, что современная психология рассматривает предметное восприятие как активный процесс, как поиск нужной информации.
[…]
Зрительный анализатор. Сенсорные зрительные расстройства
Человек […] основную информацию о внешнем мире он получает через зрительные каналы. Поэтому роль зрительного анализатора для психических функций человека трудно переоценить.
Зрительный анализатор, как и все анализаторные системы, организован по иерархическому принципу.
Основными уровнями зрительной системы каждого полушария являются: сетчатка глаза (периферический уровень); зрительный нерв (II пара); область пересечения зрительных нервов (хиазма); зрительный канатик
(место выхода зрительного пути из области хиазмы); наружное или латеральное коленчатое тело (НКТ или ЛКТ); подушка зрительного бугра, где заканчиваются некоторые волокна зрительного пути; путь от наружного коленчатого тела к коре (зрительное сияние) и первичное 17-е поле коры мозга. Работа зрительной системы обеспечивается II, III, IV и VI парами черепно-мозговых нервов.
Поражение каждого из перечисленных уровней, или звеньев, зрительной системы характеризуется особыми зрительными симптомами, особыми нарушениями зрительных функций.
Первый уровень зрительной системы — сетчатка глаза — представляет собой очень сложный орган, который называют «куском мозга, вынесенным наружу».
Рецепторный строй сетчатки содержит два типа рецепторов:
♦ колбочки (аппарат дневного, фотопического зрения);
♦ палочки (аппарат сумеречного, скотопического зрения).
Когда свет достигает глаза, возникающая в этих элементах фотопическая реакция преобразуется в импульсы, передающиеся через различные уровни зрительной системы в первичную зрительную кору (17-е поле). Количество колбочек и палочек неравномерно распределено в разных областях сетчатки; колбочек значительно больше в центральной части сетчатки (fovea) — зоне максимально ясного зрения.
Эта зона несколько сдвинута в сторону от места выхода зрительного нерва — области, которая называется слепым пятном (papilla n. optici).
Человек относится к числу так называемых фронтальных млекопитающих, т. е. животных, у которых глаза расположены во фронтальной плоскости. Вследствие этого зрительные поля обоих глаз (т. е. та часть зрительной среды, которая воспринимается каждой сетчаткой отдельно) перекрываются. Это перекрытие зрительных полей является очень важным эволюционным приобретением, позволившим человеку выполнять точные манипуляции руками под контролем зрения, а также обеспечившим точность и глубину видения (бинокулярное зрение). Благодаря бинокулярному зрению появилась возможность совмещать образы объекта, возникающие в сетчатках обоих глаз, что резко улучшило восприятие глубины изображения, его пространственных признаков.
Зона перекрытия зрительных полей обоих глаз составляет приблизительно 120°. Зона монокулярного видения составляет около 30° для каждого глаза; эту зону мы видим только одним глазом, если фиксировать центральную точку общего для двух глаз поля зрения.
Зрительная информация, воспринимаемая двумя глазами или только одним глазом (левым или правым), проецируется на разные отделы сетчатки и, следовательно, поступает в разные звенья зрительной системы.
В целом, участки сетчатки, расположенные к носу от средней линии (нозальные отделы), участвуют в механизмах бинокулярного зрения, а участки, расположенные в височных отделах (темпоральные отделы), — в монокулярном зрении.
Кроме того, важно помнить, что сетчатка организована и по верхненижнему принципу: ее верхние и нижние отделы представлены на разных уровнях зрительной системы по-разному. Знания об этих особенностях строения сетчатки позволяют диагностировать ее заболевания.
Поражения сетчаточного уровня зрительной системы разнообразны: это разные формы дегенерации сетчатки; кровоизлияния; различные заболевания глаз, в которых поражается также и сетчатка (центральное место среди этих поражений занимает такое распространенное заболевание, как глаукома). Во всех этих случаях поражение, как правило, одностороннее, т. е. зрение нарушается только в одном глазу; далее — это относительно элементарное расстройство остроты зрения (т. е. остроты светоощущения), или полей зрения (по типу скотомы), или цветоощущения. Зрительные функции второго глаза остаются сохранными. Отсутствуют и более сложные зрительные расстройства.
Второй уровень работы зрительной системы — зрительные нервы (II пара). Они очень коротки и расположены сзади глазных яблок в передней черепной ямке, на базальной поверхности больших полушарий головного мозга. Разные волокна зрительных нервов несут зрительную информацию от разных отделов сетчаток. Волокна от внутренних участков сетчаток проходят во внутренней части зрительного нерва, от наружных участков — в наружной, от верхних участков — в верхней, а от нижних — в нижней. Поражения зрительного нерва встречаются в клинике локальных поражений головного мозга довольно часто в связи с различными патологическими процессами в передней черепной ямке: опухолями, кровоизлияниями, воспалительными процессами и др. Такое поражение зрительного нерва приводит к расстройству сенсорных зрительных функций только в одном глазу, причем в зависимости от места поражения страдают зрительные функции соответствующих участков сетчатки. Важным симптомом поражения зрительного нерва является отек начала (соска) зрительного нерва (слева или справа), который может привести к его атрофии.
Область хиазмы составляет третье звено зрительной системы. Как известно, у человека в зоне хиазмы происходит неполный перекрест зрительных путей. Волокна от нозальных половин сетчаток поступают в противоположное (контралатеральное) полушарие, а волокна от темпоральных половин — в ипсилатеральное. Благодаря неполному перекресту зрительных путей зрительная информация от каждого глаза поступает в оба полушария. Важно помнить, что волокна, идущие от верхних отделов сетчаток обоих глаз, образуют верхнюю половину хиазмы, а идущие от нижних отделов — нижнюю; волокна от fovea также подвергаются частичному перекресту и расположены в центре хиазмы. При поражении хиазмы возникают различные (чаще симметричные) нарушения полей зрения обоих глаз (гемианопсии) вследствие поражения соответствующих волокон, идущих от сетчаток. Поражение разных отделов хиазмы приводит к появлению разных видов гемианопсий:
♦ битемпоральной;
♦ бинозальной;
♦ верхней квадрантной;
♦ нижней квадрантной;
♦ односторонней нозальной гемианопсии (при разрушении наружной части хиазмы с одной стороны).
Гемианопсия может быть полной или частичной; в последнем случае возникают скотомы (частичное
выпадение) в соответствующих отделах полей зрения. Все перечисленные виды гемианопсий характерны только для поражения хиазмального уровня зрительной системы.
При поражении зрительных канатиков (fractes opticus), соединяющих область хиазмы с наружным коленчатым телом, возникает гомонимная (односторонняя) гемианопсия, сторона которой определяется стороной поражения. Гомонимные гемианопсии могут быть полными или неполными. Особенностью этого типа гемианопсий является то, что вследствие поражения волокон, идущих от области fovea, граница между пораженным и сохранным полями зрения проходит в виде вертикальной линии.
Четвертый уровень зрительной системы — наружное или латеральное коленчатое тело (НКТ или ЛКТ).
Это часть зрительного бугра, важнейшее из таламических ядер, представляет собой крупное образование, состоящее из нервных клеток, где сосредоточен второй нейрон зрительного пути (первый нейрон находится в сетчатке). Таким образом, зрительная информация без какой-либо переработки поступает непосредственно из сетчатки в НКТ. У человека 80 % зрительных путей, идущих от сетчатки, заканчиваются в НКТ, остальные 20 % идут в другие образования (подушку зрительного бугра, переднее двухолмие, стволовую часть мозга), что указывает на высокий уровень кортикализации зрительных функций.
НКТ, как и сетчатка, характеризуется топическим строением, т. е. различным областям сетчатки соответствуют различные группы нервных клеток в НКТ. Кроме того, в разных участках НКТ представлены области зрительного поля, которые воспринимаются одним глазом (зоны монокулярного видения), и области, которые воспринимаются двумя глазами (зоны бинокулярного видения), а также область центрального видения. При полном поражении НКТ возникает полная односторонняя гемианопсия (левосторонняя или правосторонняя), при частичном поражении — неполная, с границей в виде вертикальной линии.
В том случае, когда очаг поражения находится рядом с НКТ и раздражает его, иногда возникают сложные синдромы в виде зрительных галлюцинаций, связанных с нарушениями сознания.
Как уже было сказано выше, помимо НКТ существуют и другие инстанции, куда поступает зрительная информация, — это подушка зрительного бугра, переднее двухолмие и стволовая часть мозга. При их поражении никаких нарушений зрительных функций как таковых не возникает, что указывает на иное их назначение. Переднее двухолмие, как известно, регулирует целый ряд двигательных рефлексов (типа старт-рефлексов), в том числе и тех, которые «запускаются» зрительной информацией. По-видимому, сходные функции выполняет и подушка зрительного бугра, связанная с большим количеством инстанций, в частности — с областью базальных ядер. Стволовые структуры мозга участвуют в регуляции общей неспецифической активации мозга через коллатерали, идущие от зрительных путей. Таким образом, зрительная информация, идущая в стволовую часть мозга, является одним из источников, поддерживающих активность неспецифической системы.
Пятый уровень зрительной системы — зрительное сияние (пучок Грациоле) — довольно протяженный участок мозга, находящийся в глубине теменной и затылочной долей. Это широкий, занимающий большое пространство веер волокон, несущих зрительную информацию от разных участков сетчатки в разные области 17-го поля коры. Эта область мозга поражается весьма часто (при кровоизлияниях, опухолях, травмах и др.), что приводит к гомонимной гемианопсии, т. е. выпадению полей зрения (левого или правого). Из-за широкого расхождения волокон в пучке Грациоле гомонимная гемианопсия часто является неполной, т. е. слепота не распространяется на всю левую (или правую) половину поля зрения. Последняя инстанция — первичное 17-е поле коры больших полушарий, расположено главным образом на медиальной поверхности мозга в виде треугольника, который направлен острием вглубь мозга. Это значительная по протяженности площадь коры больших полушарий по сравнению с первичными корковыми полями других анализаторов, что отражает роль зрения в жизни человека. Важнейшим анатомическим признаком 17-го поля является хорошее развитие IV слоя коры, куда приходят зрительные афферентные импульсы; IV слой связан с V слоем, откуда «запускаются» местные двигательные рефлексы, что характеризует «первичный нейронный комплекс коры».
17-е поле организовано по топическому принципу, т. е. разные области сетчатки представлены в его разных участках. Это поле имеет две координаты: верхне-нижнюю и передне-заднюю. Верхняя часть 17-го поля связана с верхней частью сетчатки, т. е. с нижними полями зрения; в нижнюю часть 17-го поля поступают импульсы от нижних участков сетчатки, т. е. от верхних полей зрения.
В задней части 17-го поля представлено бинокулярное зрение в передней части — периферическое монокулярное зрение.
При поражении 17-го поля в левом и правом полушариях одновременно (что может быть, например, при ранениях затылочного полюса) возникает центральная слепота. Когда же поражение захватывает 17-е поле одного полушария, возникает выпадение полей зрения с одной стороны, причем при правостороннем очаге возможна «фиксированная» левосторонняя гемианопсия, когда больной как бы не замечает своего зрительного дефекта. При поражении 17-го поля граница между «хорошим» и «плохим» участками полей зрения проходит не в виде вертикальной линии, а в виде полукруга в зоне fovea, так как при этом сохраняется область центрального видения, которая у человека представлена в обоих полушариях, что и определяет контур пограничной линии. Эта особенность позволяет различать корковую и подкорковые гемианопсии.
Как правило, у больных имеется не полное, а лишь частичное поражение 17-го поля, что приводит к частичному выпадению полей зрения (скотомам); при этом участки нарушенных полей зрения по форме и величине в обоих глазах симметричны. При менее грубых поражениях 17-го поля возникают частичные нарушения зрительных функций в виде снижения (изменения) цветоощущения, фотопсий (т. е. ощущение ярких вспышек, «искр», иногда окрашенных, появляющихся в определенном участке поля зрения). Все описанные выше нарушения зрительных функций относятся к сенсорным, относительно элементарным нарушениям, которые непосредственно не связаны с высшими зрительными функциями, хотя и являются их основой.
Гностические зрительные расстройства
Высшие гностические зрительные функции обеспечиваются прежде всего работой вторичных полей зрительной системы (18-е и 19-е) и прилегающих к ним третичных полей коры больших полушарий. Вторичные 18-е и 19-е поля расположены как на наружной конвекситальной, так и на внутренней медиальной поверхностях больших полушарий. Они характеризуются хорошо развитым III слоем, в котором осуществляется переключение импульсов из одного участка коры в другой.
При электрическом раздражении 18-го и 19-го полей возникает не локальное, точечное возбуждение, как при раздражении 17-го поля, а активация широкой зоны, что свидетельствует о широких ассоциативных связях этих областей коры.
Из исследований, проведенных на человеке У. Пенфилдом, Г. Джаспером (1959) и рядом других авторов, известно, что при электрическом раздражении 18-го и 19-го полей появляются сложные зрительные образы. Это уже не отдельные вспышки света, а знакомые лица, картины, иногда какие-то неопределенные образы. Основные сведения о роли этих участков коры больших полушарий в зрительных функциях получены из клиники локальных поражений головного мозга. Клинические наблюдения показывают, что поражение этих областей коры и прилегающих к ним подкорковых зон («ближайшей подкорки», по выражению А.Р. Лурия) приводит к различным нарушениям зрительного гнозиса. Эти нарушения получили название зрительные агнозии. Этим термином обозначаются расстройства зрительного восприятия, возникающие при поражении корковых структур задних отделов больших полушарий и протекающие при относительной сохранности элементарных зрительных функций (остроты зрения, полей зрения, цветоощущения). При всех формах агностических зрительных расстройств элементарные сенсорные зрительные функции остаются относительно сохранными, т. е. больные достаточно хорошо видят, у них нормальное цветоощущение, часто сохранны и поля зрения; иными словами, у них как будто бы есть все предпосылки, чтобы воспринимать объекты правильно. Однако у них нарушен именно гностический уровень работы зрительной системы. В некоторых случаях у больных, помимо гностических, имеются нарушения и сенсорных функций. Но это, как правило, относительно тонкие дефекты, которые не могут объяснить выраженность и характер нарушений высших зрительных функций.
[…] В клинике локальных поражений головного мозга описаны разнообразные формы нарушений высших зрительных функций, или разные формы зрительных агнозий. […]
Большинство авторов, исходя из клинической феноменологии, выделяет шесть основных форм нарушений зрительного гнозиса:
1)если больной, правильно оценивая отдельные элементы объекта (или его изображения), не может понять его смысла в целом — это называется предметной агнозией;
2) если он не различает человеческие лица (или фотографии) — лицевой агнозией;
3)если он плохо ориентируется в пространственных признаках изображения — оптико-пространственной агнозией;
4) если он, правильно копируя буквы, не может их читать — буквенной агнозией;
5) если он различает цвета, но не знает, какие предметы окрашены в данный цвет, т. е. не может вспомнить цвет знакомых предметов, — цветовой агнозией;
6) как самостоятельная форма выделяется и симультанная агнозия — такое нарушение зрительного гнозиса, когда больной может воспринимать только отдельные фрагменты изображения, причем этот дефект наблюдается и при сохранности полей зрения.
[…]
Клинические наблюдения показывают, что форма нарушений зрительного гнозиса связана как со стороной поражения мозга, так и с локализацией поражения внутри «широкой зрительной сферы» — конвекситальной коры затылочных и теменных отделов мозга, где выделяют две основные подобласти: нижнюю и верхнюю части.
Рассмотрим несколько подробнее разные формы зрительных агнозий.
Предметная агнозия — одна из самых распространенных форм нарушений зрительного гнозиса, которая в той или иной степени встречается у большинства больных с поражением затылочно-теменных отделов мозга. В грубой форме предметная агнозия наблюдается лишь при двухстороннем поражении затылочно-теменных отделов мозга, т. е. при двухстороннем поражении 18-го и 19-го полей. Предметная зрительная агнозия связана с поражением нижней части «широкой зрительной сферы». Она характеризуется тем, что больной видит как будто бы все, он может описать отдельные признаки предмета, но не может сказать, что же это такое. Особо грубое нарушение возможности правильно оценить предмет возникает при двухстороннем поражении нижних частей широкой зрительной сферы: больной, глядя на предмет, не может его опознать, но при его ощупывании часто правильно решает эту задачу. В своей повседневной жизни такие больные ведут себя почти как слепые и хотя они не натыкаются на предметы, но постоянно ощупывают их или ориентируются по звукам. Однако в такой грубой форме предметная агнозия встречается сравнительно редко, чаще она проявляется в скрытой форме при выполнении специальных зрительных задач: например, при распознавании контурных, перечеркнутых, наложенных друг на друга, перевернутых изображений и т. д.
Так, при совмещении 3, 4, 5 контуров (проба Поппельрейтера) здоровый человек видит контуры всех объектов; у больных эта задача вызывает большие трудности: они не могут выделить отдельные контуры и видят просто путаницу линий.
При предметной агнозии трудности опознания формы объектов являются первичными, и в наиболее «чистом» виде они проявляются при опознании именно контуров объектов; в то же время копирование рисунков у них может быть сохранным.
У больных с предметной агнозией (как и с другими формами нарушений зрительного гнозиса) грубо изменяются временные характеристики зрительного восприятия. Тахистоскопическими исследованиями установлено, что у таких больных резко увеличиваются пороги узнавания изображений; причем, как правило, они возрастают на несколько порядков. Если здоровый человек воспринимает простые изображения за 5—10 мс (без фонового стирающего изображения), то у больных время опознания простых изображений возрастает до 1с. и больше. Таким образом, при зрительных агнозиях наблюдается совсем другой режим работы зрительной системы, что и вызывает большие сложности в переработке зрительной информации.
Оптико-пространственная агнозия связана преимущественно с поражением верхней части «широкой зрительной сферы». В особенно грубой форме она наблюдается при двухстороннем поражении затылочно-теменных отделов мозга. Однако и при одностороннем поражении эти нарушения также выражены достаточно отчетливо.
При оптико-пространственной агнозии у больных теряется возможность ориентации в пространственных признаках окружающей среды и изображений объектов. У них нарушается лево-правая ориентировка; они перестают понимать ту символику рисунка, которая отражает пространственные признаки объектов. Такие больные не понимают географическую карту, у них нарушена ориентировка в странах света. Описанию подобного нарушения посвящена книга А.Р. Лурия «Потерянный и возвращенный мир» (1971), где рассказывается о больном, в прошлом топографе, получившем ранение затылочно-теменной области левого полушария мозга.
В грубых случаях у больных нарушается ориентировка не только в лево-правых, но и в верхне-нижних координатах. У больных с оптико-пространственной (как и с предметной) агнозией, как правило, нарушается способность рисовать (при относительной сохранности способности копировать изображение). Они не умеют передавать на рисунке пространственные признаки объектов (дальше-ближе, больше-меньше, слева-справа, сверху-снизу). В некоторых случаях распадается даже общая схема рисунка. Так, больные, рисуя человека, отдельно изображают части его тела (руки, ноги, глаза, нос и т. д.) и не знают, как их соединить. Чаще нарушается рисунок при поражении задних отделов правого полушария. В ряде случаев (как правило, при правополушарных очагах) наблюдается односторонняя оптико-пространственная агнозия, когда больные, даже копируя рисунок, изображают только одну сторону предмета или грубо искажают изображение одной (чаще левой) стороны.
Одновременно у них часто нарушается также и возможность зрительной афферентации пространственно-организованных движений, т. е. «праксис позы». Такие больные не могут скопировать позу, показанную им экспериментатором; не знают, как расположить руку по отношению к своему телу; у них отсутствует та непосредственная легкость восприятия пространственных отношений, которая присуща здоровым людям, и это затрудняет копирование поз по зрительному образцу (выполняемых одной или двумя руками).

Рисунок: рисунки больных о оптико-пространственной агнозией:
А — рисунки куба больными с поражением правой затылочно-теменной области мозга;Б — рисунки больных с поражением левой теменной области мозга
С этим связаны различные трудности в бытовых двигательных актах, в которых требуется пространственная ориентация движений. Эти больные плохо выполняют движения, требующие элементарной зрительно-пространственной ориентировки, например не могут постелить покрывало на постель, надеть пиджак, брюки и т. п. Подобные нарушения получили название «апраксии одевания». Сочетания зрительно-пространственных и двигательно-пространственных расстройств называют «апрактоагнозией».

Рисунок: копирование рисунков больным с поражением задних отделов правого полушария, имеющим одностороннюю оптико-пространственную агнозию.
Больной игнорирует левую сторону рисунков
Оптико-пространственные нарушения влияют иногда и на навык чтения. В этих случаях возникают трудности прочтения таких букв, которые имеют «лево-правые» признаки, Больные не могут различить правильно и неверно написанные буквы (например: К, М, Р,Ч и др.), и это задание может быть одним из тестов на определение зрительной ориентировки в пространственных признаках объектов. В подобных случаях нарушения опознания букв с зеркальными пространственными признаками, как правило, отражают общий дефект пространственной ориентировки в объектах.
Особой формой зрительных агностических расстройств является буквенная агнозия. В чистом виде буквенная агнозия проявляется в том, что больные, совершенно правильно копируя буквы, не могут их назвать. У них распадается навык чтения (первичная алексия).
Подобное нарушение чтения встречается изолированно от других нарушений высших зрительных функций, что и дает основание выделить этот дефект в самостоятельную форму агнозии. Такие больные правильно воспринимают предметы, правильно оценивают их изображения и даже правильно ориентируются в сложных пространственных изображениях и реальных объектах, однако они «не понимают» буквы и не могут читать.
Такая форма агнозии, как правило, встречается при поражении левого полушария мозга — нижней части «широкой зрительной сферы» (у правшей).
Цветовая агнозия также представляет собой самостоятельный тип зрительных гностических расстройств. Различают собственно цветовую агнозию и нарушение распознавания цветов как таковых (цветовая слепота или дефекты цветоощущений). Цветовая слепота и нарушение цветоощущений могут иметь как периферическое, так и центральное происхождение, т. е. быть связанными с поражением как сетчатки, так и подкорковых и коркового звеньев зрительной системы. Известно, что цветоощущение возникает при действии трех различных типов колбочек (сетчаточных детекторов), которые чувствительны к различным цветам: сине-зеленым, красно-зеленым и желтым. Эта способность колбочек быть реактивными на определенные цветовые раздражения является основой цветоощущения, и дефект этой способности может быть вызван разного типа поражениями сетчатки (дегенерацией и т. п.).
Известны нарушения цветоразличения, связанные с поражением НКТ и затылочной коры (17-го поля), что указывает на существование в зрительной системе специального канала (или каналов), предназначенного для проведения информации о цвете объекта.
Цветовая агнозия, в отличие от нарушений цветоразличения, является нарушением высших зрительных функций. В клинике описаны нарушения цветового гнозиса, которые наблюдаются на фоне сохранного цветоощущения. Такие больные правильно различают отдельные цвета и правильно их называют. Однако им трудно, например, соотнести цвет с определенным предметом и наоборот; они не могут вспомнить, каков цвет апельсина, моркови, елки и т. д. Больные не могут назвать предметы определенного конкретного цвета. У них отсутствует обобщенное представление о цвете, и поэтому они не в состоянии выполнить процедуру классификации цветов, что связано не с трудностями различения цветов, а с трудностями их категоризации. Известно, что человек воспринимает огромное количество оттенков цветов, но названий цветов (категорий) сравнительно мало. Поэтому в обычной жизни здоровый человек постоянно решает задачу на категоризацию цвета. Именно эта категоризация цветовых ощущений затруднена у больных с цветовой агнозией.
Особую форму зрительной агнозии составляет симультанная агнозия. Долгое время она была известна как синдром Балинта. Эта форма нарушения зрительного гнозиса проявляется в том, что больной одновременно не может воспринимать двух изображений, так как у него резко сужен объем зрительного восприятия.
Больной не может воспринять целое, он видит только его часть (или части).
Возникает вопрос: почему больной не может перевести взор и рассмотреть все изображение последовательно? Это происходит потому, что синдром Балинта всегда сопровождается сложными нарушениями движений глаз, которые называются «атаксия взора».
Взор больного становится неуправляемым, глаза совершают непроизвольные скачки, постоянно находясь в движении. Это создает трудности в организованном зрительном поиске, вследствие чего больной не может рассматривать объект последовательно. Предполагается, что причиной симультанной агнозии является слабость корковых зрительных клеток, которые способны лишь на узколокальные очаги возбуждения. Связь синдрома Балинта со стороной поражения и локализацией очага внутри «широкой зрительной сферы» пока не установлена.
Лицевая агнозия — особая форма нарушений зрительного гнозиса, которая проявляется в том, что у больного теряется способность распознавать реальные лица или их изображения (на фотографиях, рисунках и т. п.).
При грубой форме лицевой агнозии больные не могут различить женские и мужские лица, а также лица детей и взрослых; не узнают лица своих родных и близких. Такие больные узнают людей (включая и самых близких) только по голосу. Лицевая агнозия четко связана с поражением задних отделов правого полушария (у правшей), в большей степени — нижних отделов «широкой зрительной сферы».
В целом вопрос о связи разных форм зрительных агностических расстройств со стороной и зоной поражения затылочно-теменных отделов мозга окончательно не решен. Многие авторы указывают, что различные формы зрительных агнозий проявляются особенно отчетливо при повреждении комиссуральных волокон валика мозолистого тела, соединяющих 18-е и 19-е поля левого и правого полушарий головного мозга.
Особый интерес для понимания механизмов зрительных агнозий представляют исследования движений глаз (регулируемых III, IV и VI парами черепно-мозговых нервов) при различных формах нарушения зрительного восприятия. Нарушения зрительного гнозиса коррелируют с различными нарушениями глазодвигательной активности, сопровождающей зрительное восприятие объекта. Это могут быть явления инактивности движений глаз, глазодвигательные персеверации, игнорирование одной стороны зрительного поля и др.
Вопрос о роли движений глаз в нарушениях зрительного гнозиса является дискуссионным. Согласно одной точке зрения движения глаз, обводящие контур объекта, — обязательный механизм зрительного восприятия. Однако, как показали исследования, многие формы зрительных агнозий протекают при сохранной глазодвигательной активности.
[…]
Смирнов В.М., Будылина С.М. Физиология сенсорных систем и высшая нервная деятельность: Учеб. пособие для студ. высш. учеб. заведений. – М.: Академия, 2003. – с. 54 – 63
Слуховой анализатор
С помощью слухового анализатора человек ориентируется в звуковых сигналах окружающей среды, формирует соответствующие поведенческие реакции, например оборонительные или пищедобывательные. Способность восприятия человеком разговорной и вокальной речи, музыкальных произведений делает слуховой анализатор необходимым компонентом средств общения, познания, приспособления.
Адекватным раздражителем для слухового анализатора являются звуки, т.е. колебательные движения частиц упругих тел, распространяющихся в виде волн в самых различных средах, включая воздушную среду, и воспринимающиеся ухом. Звуковые волновые колебания (звуковые волны) характеризуются чистотой и амплитудой. Частота звуковых волн определяет высоту звука. Человек различает звуковые волны с частотой от 20 до 20 000 Гц. Звуки, частота которых ниже 20 Гц — инфразвуки и выше 20 000 Гц , (20 кГц) — ультразвуки, человеком не ощущаются. Звуковые волны, имеющие синусоидальные, или гармонические, колебания, называют тоном. Звук, состоящий из не связанных между собой, частот, называют шумом. При большой частоте звуковых волн — тон высокий, при малой — низкий.
Второй характеристикой звука, которую различает слуховая сенсорная система, является его сила, зависящая от амплитуды звуковых волн. Сила звука или его интенсивность воспринимаются человеком как громкость. Ощущение громкости нарастает при усилении звука и зависит также от частоты звуковых колебаний, т. е. громкость звучания определяется взаимодействием интенсивности (силы) и высоты (частоты) звука. Единицей измерения громкости звука является бел, в практике обычно используется децибел (дБ), т.е. 0,1 бела. Человек различает звуки также по тембру, или «окраске». Тембр звукового сигнала зависит от спектра, т.е. от состава дополнительных частот (обертонов), которые сопровождают основной тон (частоту). По тембру можно различить звуки одинаковой высоты и громкости, на чем основано узнавание людей по голосу. Чувствительность слухового анализатора определяется минимальной силой звука, достаточной для возникновения слухового ощущения. В области звуковых колебаний от 1000 до 3000 в секунду, что соответствует человеческой речи, ухо обладает наибольшей чувствительностью. Эта совокупность частот получила название речевой зоны. В данной области воспринимаются звуки, имеющие давление меньше чем 0,001 бара (1 бар составляет приблизительно одну миллионную часть нормального атмосферного давления). Исходя из этого, в передающих устройствах, чтобы обеспечить адекватное понимание речи, речевая информация должна передаваться в речевом диапазоне частот.
Структурно-функциональная характеристика
Рецепторный (периферический) отдел слухового анализатора, превращающий энергию звуковых волн в энергию нервного возбуждения, представлен рецепторными волосковыми клетками кортиева органа (орган Корти), находящимися в улитке.

Рисунок: Орган слуха
Слуховые рецепторы (фонорецепторы) относятся к механорецепторам, являются вторичными и представлены внутренними и наружными волосковыми клетками. У человека приблизительно 3500 внутренних и 20 000 наружных волосковых клеток, которые расположены на основной мембране внутри среднего канала внутреннего уха.
Внутреннее ухо (звуковоспринимающий аппарат), а также среднее ухо (звукопередающий аппарат) и наружное ухо (звукоулавливающий аппарат) объединяются в понятие орган слуха.
Наружное ухо за счет ушной раковины обеспечивает улавливание звуков, концентрацию их в направлении наружного слухового прохода и усиление интенсивности звуков. Кроме того, структуры наружного уха выполняют защитную функцию, охраняя барабанную перепонку от механических и температурных воздействий внешней среды.
Среднее ухо (звукопроводящий отдел) представлено барабанной полостью, где расположены три слуховые косточки: молоточек, наковальня и стремечко. От наружного слухового прохода среднее ухо отделено барабанной перепонкой. Рукоятка молоточка вплетена в барабанную перепонку, другой его конец сочленен с наковальней, которая, в свою очередь, сочленена со стремечком. Стремечко прилегает к мембране овального окна. Площадь барабанной перепонки (70 мм2) значительно больше площади овального окна (3,2 мм2), благодаря чему происходит усиление давления звуковых волн на мембрану овального окна примерно в 25 раз. Так как рычажный механизм косточек уменьшает амплитуду звуковых волн примерно в 2 раза, то, следовательно, происходит такое же усиление звуковых волн на овальном окне. Таким образом, происходит общее усиление звука средним ухом примерно в 60 – 70 раз. Если же учитывать усиливающий эффект наружного уха, то эта величина достигает 180 – 200 раз. Среднее ухо имеет специальный защитный механизм, представленный двумя мышцами: мышцей, натягивающей барабанную перепонку, и мышцей, фиксирующей стремечко. Степень сокращения этих мышц зависит от силы звуковых колебаний. При сильных звуковых колебаниях мышцы ограничивают амплитуду колебаний барабанной перепонки и движение стремечка, предохраняя тем самым рецепторный аппарат во внутреннем ухе от чрезмерного возбуждения и разрушения. При мгновенных сильных раздражениях (удар в колокол) этот защитный механизм не успевает срабатывать. Сокращение обеих мышц барабанной полости осуществляется по механизму безусловного рефлекса, который замыкается на уровне стволовых отделов мозга. В барабанной полости поддерживается давление, равное атмосферному, что очень важно для адекватного восприятия звуков. Эту функцию выполняет евстахиева труба, которая соединяет полость среднего уха с глоткой. При глотании труба открывается, вентилируя полость среднего уха и уравнивая давление в нем с атмосферным. Если внешнее давление быстро меняется (быстрый подъем на высоту), а глотания не происходит, то разность давлений между атмосферным воздухом и воздухом в барабанной полости приводит к натяжению барабанной перепонки и возникновению неприятных ощущений, снижению восприятия звуков.

Рисунок: кортиев орган
Внутреннее ухо представлено улиткой — спирально закрученным костным каналом, имеющим 2,5 завитка, который разделен основной мембраной и мембраной Рейснера на три узких части (лестницы). Верхний канал (вестибулярная лестница) начинается от овального окна и соединяется с нижним каналом (барабанной лестницей) через геликотрему (отверстие в верхушке) и заканчивается круглым окном. Оба канала представляют собой единое целое и заполнены перилимфой, сходной по составу со спинномозговой жидкостью. Между верхним и нижним каналами находится средний (средняя лестница). Он изолирован и заполнен эндолимфой. Внутри среднего канала на основной мембране расположен собственно звуковоспринимающий аппарат — орган Корти (кортиев орган) с рецепторными клетками, представляющий периферический отдел слухового анализатора.
Основная мембрана вблизи овального окна по ширине составляет 0,04 мм, затем по направлению к вершине она постепенно расширяется, достигая у геликотремы 0,5 мм. Над кортиевым органом лежит текториалъная (покровная) мембрана соединительнотканного происхождения, один край которой закреплен, второй — свободен. Волоски наружных и внутренних волосковых клеток соприкасаются с текториальной мембраной. При этом изменяется проводимость ионных каналов рецепторных (волосковых) клеток, формируются микрофонный и сум-мационный рецепторные потенциалы. Образуется и выделяется медиатор ацетилхолин в синаптическую щель рецепторно-афферентного синапса. Все это приводит к возбуждению волокна слухового нерва, к возникновению в нем потенциала действия. Так происходит трансформация энергии звуковых волн в нервный импульс. Каждое волокно слухового нерва имеет кривую частотной настройки, которая называется также частотно-пороговой кривой. Этот показатель характеризует площадь рецептивного поля волокна, которая может быть узкой или широкой. Узкой она бывает при тихих звуках, а при увеличении их интенсивности расширяется.
Проводниковый отдел слухового анализатора представлен периферическим биполярным нейроном, расположенным в спиральном ганглии улитки (первый нейрон). Волокна слухового (или кохлеарного) нерва, образованные аксонами нейронов спирального ганглия, заканчиваются на клетках ядер кохлеарного комплекса продолговатого мозга (второй нейрон). Затем после частичного перекреста волокна идут в медиальное коленчатое тело метаталамуса, где опять происходит переключение (третий нейрон), отсюда возбуждение поступает в кору (четвертый нейрон). В медиальных (внутренних) коленчатых телах, а также в нижних буграх четверохолмия располагаются центры рефлекторных двигательных реакций, возникающих при действии звука.
Центральный, или корковый, отдел слухового анализатора находится в верхней части височной доли большого мозга (верхняя височная извилина, поля 41 и 42 по Бродману). Важное значение для функции слухового анализатора имеют поперечные височные извилины (извилины Гешля).
Слуховая сенсорная система дополняется механизмами обратной связи, обеспечивающими регуляцию деятельности всех уровней слухового анализатора с участием нисходящих путей. Такие пути начинаются от клеток слуховой коры, переключаясь последовательно в медиальных коленчатых телах метаталамуса, задних (нижних) буграх четверохолмия, в ядрах кохлеарного комплекса. Входя в состав слухового нерва, центробежные волокна достигают волосковых клеток кортиева органа и настраивают их на восприятие определенных звуковых сигналов.
Восприятие высоты, силы звука и локализации источника звука
Восприятие высоты, силы звука и локализации источника звука начинается с попадания звуковых волн в наружное ухо, где они приводят в движение барабанную перепонку. Колебания барабанной перепонки через систему слуховых косточек среднего уха передаются на мембрану овального окна, что вызывает колебание перилимфы вестибулярной (верхней) лестницы. Эти колебания через геликотрему передаются перилимфе барабанной (нижней) лестницы и доходят до круглого окна, смещая его мембрану по направлению к полости среднего уха.
Колебания перилимфы передаются также на эндолимфу перепончатого (среднего) канала, что приводит в колебательные движения основную мембрану, состоящую из отдельных волокон, натянутых, как струны рояля. При действии звука волокна мембраны приходят в колебательные движения вместе с рецепторными клетками кортиева органа, расположенными на них. При этом волоски рецепторных клеток контактируют с текториальной мембраной, реснички волосковых клеток деформируются. Возникает вначале рецепторный потенциал, а затем потенциал действия (нервный импульс), который далее проводится по слуховому нерву и передается в другие отделы слухового анализатора.
Восприятие звуков различной высоты (частоты), согласно резонансной теории Гельмгольца, обусловлено тем, что каждое волокно основной мембраны настроено на звук определенной частоты. Так, звуки низкой частоты воспринимаются длинными волнами основной мембраны, расположенными ближе к верхушке улитки, звуки высокой частоты воспринимаются короткими волокнами основной мембраны, расположенными ближе к основанию улитки. При действии сложного звука возникают колебания различных волокон мембраны.
В современной интерпретации резонансный механизм лежит в основе теории места, в соответствии с которой в состояние колебания вступает вся мембрана. Однако максимальное отклонение основной мембраны улитки происходит только в определенном месте. При увеличении частоты звуковых колебаний максимальное отклонение основной мембраны смещается к основанию улитки, где располагаются более короткие волокна основной мембраны, — у коротких волокон возможна более высокая частота колебаний. Возбуждение волосковых клеток именно этого участка мембраны при посредстве медиатора передается на волокна слухового нерва в виде определенного числа импульсов, частота следования которых ниже частоты звуковых волн (лабильность нервных волокон не превышает 800 —1000 Гц). Частота воспринимаемых звуковых волн достигает 20 000 Гц. Таким способом осуществляется пространственный тип кодирования высоты и частоты звуковых сигналов. При действии тонов примерно до 800 Гц кроме пространственного кодирования происходит еще и временное (частотное) кодирование, при котором информация передается также по определенным волокнам слухового нерва, но в виде импульсов (залпов), частота следования которых повторяет частоту звуковых колебаний. Отдельные нейроны на разных уровнях слуховой сенсорной системы настроены на определенную частоту звука, т.е. каждый нейрон имеет свой специфический частотный порог, свою определенную частоту звука, на которую реакция нейрона максимальна. Таким образом, каждый нейрон из всей совокупности звуков воспринимает лишь определенные достаточно узкие участки частотного диапазона, не совпадающие между собой, а совокупности нейронов воспринимают весь частотный диапазон слышимых звуков, что и обеспечивает полноценное слуховое восприятие.
Правомерность этого положения подтверждается результатами протезирования слуха человека, когда электроды вживлялись в слуховой нерв, а его волокна раздражались электрическими импульсами разных частот, которые соответствовали звукосочетаниям определенных слов и фраз, обеспечивая смысловое восприятие речи.
Анализ интенсивности звука также осуществляется в слуховой сенсорной системе. При этом сила звука кодируется как частотой импульсов, так и числом возбужденных рецепторов и соответствующих нейронов. В частности, наружные и внутренние волосковые рецепторные клетки имеют разные пороги возбуждения. Внутренние клетки возбуждаются при большей силе звука, чем наружные. Кроме того, у внутренних клеток пороги возбуждения также различны. В связи с этим в зависимости от интенсивности звука меняются соотношение возбужденных рецепторных клеток кортиева органа и характер импульсации, поступающей в ЦНС. Нейроны слуховой сенсорной системы имеют различные пороги реакций. При слабом звуковом сигнале в реакцию вовлекается лишь небольшое число более возбудимых нейронов, а при усилении звука возбуждаются нейроны с меньшей возбудимостью.
Необходимо отметить, что кроме воздушной проводимости имеется костная проводимость звука, т. е. проведение звука непосредственно через кости черепа. При этом звуковые колебания вызывают вибрацию костей черепа и лабиринта, что приводит к повышению давления перилимфы в вестибулярном канале больше, чем в барабанном, так как перепонка, закрывающая круглое окно, эластична, а овальное окно закрыто стремечком. В результате этого происходит смещение основной мембраны, так же как и при воздушной передаче звуковых колебаний.
Определение локализации источника звука возможно с помощью бинаурального слуха, т. е. способности слышать одновременно двумя ушами. Благодаря бинауральному слуху человек способен более точно локализовать источник звука, чем при монауральном слухе, и определять направление звука. Для высоких звуков определение их источника обусловлено разницей силы звука, поступающего к обоим ушам, вследствие различной их удаленности от источника звука. Для низких звуков важной является разность во времени между приходом одинаковых фаз звуковой волны к обоим ушам. Определение местоположения звучащего объекта осуществляется либо путем восприятия звуков непосредственно от звучащего объекта — первичная локализация, либо путем восприятия отраженных от объекта звуковых волн — вторичная локализация, или эхолокация. При помощи эхолокации ориентируются в пространстве некоторые животные (дельфины, летучие мыши). Слуховая адаптация — это изменение слуховой чувствительности в процессе действия звука. Она складывается из соответствующих изменений функционального состояния всех отделов слухового анализатора. Ухо, адаптированное к тишине, обладает более высокой чувствительностью к звуковым раздражениям (слуховая сенситизация). При длительном слушании слуховая чувствительность снижается. Большую роль в слуховой адаптации играет ретикулярная формация, которая не только изменяет активность проводникового и коркового отделов слухового анализатора, но и за счет центробежных влияний регулирует чувствительность слуховых рецепторов, определяя уровень их «настройки» на восприятие слуховых раздражителей.
Психофизиология. Под ред. Ю.И. Александрова. – СПб.: Питер, 2003
Слуховая система
В связи с возникновением речи как средства межличностного общения, слух у человека играет особую роль. Акустические (звуковые) сигналы представляют собой колебания воздуха с разной частотой и силой. Они возбуждают слуховые рецепторы, находящиеся в улитке внутреннего уха. Рецепторы активируют первые слуховые нейроны, после чего сенсорная информация передается в слуховую область коры мозга через ряд последовательных отделов, которых особенно много в слуховой системе.
Структура и функции наружного и среднего уха
Наружный слуховой проход проводит звуковые колебания к барабанной перепонке, отделяющей наружное ухо от барабанной полости, или среднего уха. Это тонкая перегородка, которая колеблется при действии звуковых колебаний, пришедших к ней через наружный слуховой проход. В среднем ухе находятся три косточки: молоточек, наковальня и стремечко, которые последовательно передают колебания барабанной перепонки во внутреннее ухо. Благодаря особенностям геометрии слуховых косточек эти колебания передаются уменьшенными в амплитуде, но увеличенными в силе. Именно поэтому даже слабые звуковые волны способны привести к колебаниям жидкости в улитке.
Структура и функции внутреннего уха
Во внутреннем ухе находится улитка, содержащая слуховые рецепторы. Улитка представляет собой костный спиральный канал, который по всей длине разделен вестибулярной и основной мембранами на три хода: верхний, средний и нижний. Полость среднего канала не сообщается с полостью других каналов и заполнена эндолимфой, а верхний и нижний каналы сообщаются друг с другом и заполнены перилимфой. Внутри среднего канала улитки на основной мембране расположен спиральный (кортиев) орган, содержащий рецепторные клетки, которые трансформируют механические колебания в электрические потенциалы.
Колебания мембраны овального окна вызывают колебания перилимфы в верхнем и нижнем каналах, кроме того, начинает колебаться и основная мембрана. На ней расположены два вида рецепторных волосковых клеток: внутренние и наружные.
Механизмы слуховой рецепции. При колебаниях основной мембраны длинные волоски рецепторных клеток касаются текторинальной мембраны и несколько наклоняются. Это приводит к натяжению тончайших нитей, которые открывают ионные каналы в мембране рецептора. Пресинаптическое окончание волосковой клетки деполяризуется, что приводит к выходу в синаптическую щель нейромедиатора (глутамата или аспартата). Воздействуя на постсинаптическую мембрану афферентного волокна, медиатор вызывает в нем генерацию возбуждающего постсинаптического потенциала и импульсов, которые распространяются в нервные центры.
Передача в мозг акустической информации. Сигналы от волосковых клеток поступают в мозг по 32 000 афферентных н. волокон, входящих в состав кохлеарной ветви восьмого черепно-мозгового нерва. Они являются дендритами ганглиозных н. клеток спирального ганглия. По волокнам слухового нерва даже в тишине следуют спонтанные импульсы с частотой до 100 имп./с. При звуковом раздражении частота импульсации в волокнах увеличивается и остается повышенной в течение всего периода, когда действует звук. Степень учащения разрядов различна у разных волокон и связана с интенсивностью и частотой звукового воздействия. В центральных отделах слуховой системы много нейронов, возбуждение которых длится в течение всего периода действия звука, а в слуховой коре разряды ряда нейронов длятся десятки секунд после его прекращения.
Анализ частоты звука (высоты тона)
При действии звуков разной частоты возбуждаются разные рецепторные клетки кортиева органа. В улитке сочетаются два типа кодирования высоты звука: пространственный и временной. Пространственное кодирование основано на определенном расположении возбужденных рецепторов на основной мембране. При действии низких и средних тонов кроме пространственного осуществляется и временное кодирование: частота следования импульсов в волокнах слухового нерва повторяет частоту звуковых колебаний. Нейроны всех уровней слуховой системы настроены на определенную частоту и интенсивность звука. Для каждого нейрона может быть найдена оптимальная частота звука, на которую порог его реакции минимален. Частотно-пороговые кривые разных клеток не совпадают, в совокупности перекрывая весь частотный диапазон слышимых звуков, что обеспечивает их полноценное восприятие.
Анализ интенсивности звука. Сила звука кодируется частотой импульсации и числом возбужденных нейронов. При слабом стимуле в реакцию вовлекается лишь небольшое количество наиболее чувствительных нейронов, а при усилении звука в реакции участвует все большее количество дополнительных нейронов с более высокими порогами.
Слуховые ощущения
Тональность (частота) звука. Человек воспринимает звуковые колебания с частотой от 16 до 20 000 Гц. Этот диапазон соответствует 10 – 11 октавам. Верхняя граница частоты воспринимаемых звуков зависит от возраста: она постепенно понижается (в старости часто не слышат высоких тонов). Различение частоты звука характеризуется тем минимальным различием по частоте двух близких звуков, которое еще улавливается человеком. При низких и средних частотах человек способен заметить различия в 1-2 Гц. Встречаются люди с абсолютным слухом: они способны точно узнавать и обозначать любой звук даже при отсутствии звука сравнения.
Слуховая чувствительность. Минимальную силу звука, слышимого человеком в половине случаев его предъявления, называют абсолютным порогом слуховой чувствительности. Пороги слышимости сильно зависят от частоты звука. В области частот от 1000 до 4000 Гц слух человека максимально чувствителен. В этих пределах слышен звук, имеющий ничтожную энергию. При звуках ниже 1000 и выше 4000 Гц чувствительность резко уменьшается: например, при 20 и при 20 000 Гц пороговая энергия звука в 1 млн раз выше. При усилении звука можно дойти до возникновения неприятного ощущения давления и даже боли в ухе. Звуки такой силы характеризуют верхний предел слышимости и ограничивают область нормального слухового восприятия. Внутри этой области лежат и так называемые речевые поля, в пределах которых распределяются звуки речи.
Громкость звука. Кажущуюся громкость звука следует отличать от его физической силы. Ощущение громкости не идет строго параллельно нарастанию интенсивности звучания.
Адаптация. Если на ухо долго действует тот или иной звук, то чувствительность к нему падает. Степень этого снижения чувствительности (адаптации) зависит от длительности, силы звука и его частоты. Участие в слуховой адаптации нейронных механизмов типа латерального и возвратного торможения несомненно. Известно также, что сокращения мышц среднего уха могут изменять энергию сигнала, передающуюся на улитку.
Бинауральный слух. Человек и животные обладают пространственным слухом, т. е. способностью определять положение источника звука в пространстве. Это свойство основано на наличии бинаурального слуха, или слушания двумя ушами. Острота бинаурального слуха у человека очень высока: положение источника звука определяется с точностью порядка 1 углового градуса. Основой этого служит способность нейронов слуховой системы оценивать различия времени прихода звука на правое и левое ухо и интенсивности звука на каждом ухе. Если источник звука находится в стороне от средней линии головы, то звуковая волна приходит на одно ухо несколько раньше и имеет большую силу, чем на другом ухе. Оценка удаленности источника звука от организма связана с ослаблением звука и изменением его тембра.
Хомская Е.Д. Нейропсихология: 4-е издание. – СПб.: Питер, 2005. – 496 с.
Сенсорные и гностические слуховые расстройства. Слуховые агнозии. Слуховой анализатор
Сенсорные слуховые расстройства
Слуховая система, или слуховой анализатор человека, — совокупность нервных структур, воспринимающих и дифференцирующих звуковые раздражения и определяющих направление и степень удаленности источника звука, т.е. осуществляющих слуховую ориентировку в пространстве. Как и все анализаторные системы, звуковой анализатор имеет уровневое строение. Основные уровни его организации:
♦ рецептор (кортиев орган улитки);
♦ слуховой нерв (VIII пара);
♦ ядра продолговатого мозга;
♦ мозжечок;
♦ средний мозг (нижние бугры четверохолмия);
♦ медиальное или внутреннее коленчатое тело (МКТ, ВКТ);
♦ слуховое сияние (пути, идущие от МКТ в кору больших полушарий);
♦ первичное поле коры (41-е поле височных долей мозга по Бродману), находящееся в извилине Гешеля.
Только из перечисления уровней слуховой системы уже видно, что она в отличие от зрительной и кожно-кинестетической систем характеризуется большим количеством звеньев. Это существенный факт, определяющий особенности работы слуховой системы. Существуют и другие анатомические особенности слухового анализатора.
Слуховая система очень древняя. Она сформировалась первоначально как система анализа вестибулярных раздражений, и только постепенно из нее выделилась специальная подсистема, занимающаяся анализом звуков. Однако принцип работы вестибулярной и слуховой систем в целом остался одним и тем же. Он основан на превращении механического колебания в нервный импульс путем воздействия эндолимфы на нервные окончания клеток, расположенных в лабиринте (часть внутреннего уха).
История возникновения слухового анализатора зафиксирована не только в общем принципе работы вестибулярной и слуховой систем, но и в тесном анатомическом единстве их организации. Как известно, периферическая часть слуховой системы находится в лабиринте — там же, где находятся и периферические рецепторы, воспринимающие вестибулярные раздражения, сигнализирующие о положении тела в пространстве. Анатомическое сходство этих двух систем состоит и в том, что VIII пара черепно-мозговых нервов, которые передают возбуждения, идущие от кортиевого органа, содержит не только слуховые волокна, но и волокна, передающие вестибулярные раздражения. Это хорошо известно из клиники, так как при поражении слухового нерва возникают как вестибулярные, так и слуховые симптомы (головокружение и одностороннее нарушение слуха).

Рисунок: схема строения слухового анализатора.
Как известно, звук характеризуется четырьмя основными физическими параметрами, которым соответствуют определенные физиологические параметры слуховых ощущений.
Первым физическим параметром звука является частота звука; ей соответствует физиологическое качество, которое определяет высоту звука.
Человеческое ухо способно воспринимать звуки широкого диапазона—от 16-20 до 16 000-20 000 Гц (по данным разных авторов). Этот разброс характеризует большие индивидуальные различия слуховой чувствительности у разных людей (в зависимости от возраста и др.). Звуки ниже 16 Гц (инфразвуки) и выше 20 000 Гц (ультразвуки) ухом человека не воспринимаются — в отличие от многих животных.
Известно, что существует зона максимальной чувствительности к определенным частотам, которая охватывает от 1000 до 3000 Гц. Это именно тот диапазон, в котором в основном происходит речевое общение людей.
Вторым физическим параметром является интенсивность звука; ей соответствует физиологический параметр — громкость звука.
Третий параметр — длительность. Он одинаково обозначается и в физических, и в физиологических единицах.
Четвертым, не менее важным параметром звуковых раздражений является звуковой спектр. Обычно звуки не являются одиночными, т. е. состоящими из одного-единственного компонента; как правило, это набор различных компонентов — тонов или обертонов (т. е. тонов, которые находятся в кратном отношении к основному тону). Весь звуковой спектр стимула определяет такой физиологический параметр, как тембр звука.
Слуховой анализатор способен не только анализировать звуки по частоте, интенсивности, длительности и тембру, т. е. выполнять непосредственно функцию анализа различных физических качеств звукового стимула, но и участвовать в ориентации в пространстве. Мы знаем, что ориентировка в пространстве — чрезвычайно сложная функция, в которой принимают участие различные анализаторные системы.
Основной системой, обеспечивающей пространственную ориентировку, является зрительная. Однако и другие анализаторы также вносят свой вклад в эту функцию.
Вклад слухового анализатора в пространственную ориентировку очень существенен, что особенно четко проявляется у слепых людей, которые хорошо ориентируются в пространстве преимущественно с помощью звуковых раздражений.
С помощью слуховой системы определяется направление звука; это означает, что звуковое пространство характеризуется такими же пространственными координатами, как и зрительное:
♦ левая-правая сторона;
♦ верх-низ;
♦ направление, угол отклонения звука от средней линии;
♦ степень удаленности источника звука от слушателя.
Две последние характеристики — направление и степень удаленности звука — также дают человеку сведения о пространственных характеристиках источника звука.
Слуховая система в отличие от других анализаторных систем имеет еще одно очень существенное свойство: на ее основе формируется человеческая речь. Поэтому внутри слуховой системы выделяют две самостоятельные подсистемы:
♦ неречевой слух, т. е. способность ориентироваться в неречевых звуках (в музыкальных тонах и шумах);
♦ речевой слух, т. е. способность слышать и анализировать звуки речи (родного или других языков).
Эти две системы имеют общие подкорковые механизмы. Однако в пределах коры больших полушарий они различаются. Это хорошо известно из нейропсихологических исследований, показавших (на материале локальных поражений головного мозга), что при поражении левой и правой височных областей коры наблюдаются различные симптомы. Речевой слух как способность к анализу звукового состава слов родного или других языков нарушается преимущественно при поражении левой височной области, а неречевой — правой (у правшей).
Речевой слух неоднороден. В нем выделяют фонематический слух, т. е. способность различать фонемы, или смыслоразличительные звуки данного языка, на которых основан звуковой анализ отдельных звуков речи, слогов и слов, и интонационные компоненты, специфические для каждого языка. Так, интонационные особенности построения английской фразы совершенно иные, чем русской. Существуют и индивидуальные особенности интонаций. С помощью интонаций передается большой объем информации: не только нормативные признаки данного языка, но и эмоциональное содержание высказывания, и, конечно, отношение самого субъекта к тому, что именно он говорит. Эта интонационная характеристика речевого сообщения имеет много общего с музыкальным слухом. Неслучайно она нарушается отдельно, независимо от фонематических особенностей речи, преимущественно при правосторонней локализации поражения мозга (у правшей).
Остановимся подробнее на неречевом слухе и его нарушениях при поражении разных уровней слуховой системы.
Как уже говорилось выше, слуховая система характеризуется большим количеством звеньев.
Слуховой путь насчитывает не менее шести нейронов, следовательно, в нем происходит значительно больше переключений, чем в других анализаторных системах.
Важно отметить также, что слуховая афферентация от одного рецептора (в отличие от зрительной и кожно-кинестетической) поступает не только в противоположное, но и в ипсилатеральное полушарие. Далее почти на всех уровнях слуховой системы (начиная с продолговатого мозга) происходит частичный перекрест слуховых путей, что обеспечивает интегративный характер слуховой афферентации. Наконец, слуховая афферентация — как и афферентации других модальностей — участвует в различных безусловных
рефлексах (рефлексах равновесия и др.).
Периферическую часть слуховой системы составляет кортиев орган, находящийся в улитке (часть внутреннего уха), откуда берет начало VIII пара черепно-мозговых нервов.
Кортиев орган представляет собой лабиринт, расположенный внутри улитки, который содержит наружные и внутренние слуховые клетки, погруженные в эндолимфу. Эти клетки являются специализированными чувствительными рецепторами, трансформирующими механические волновые колебания в электрические сигналы. При звуковых колебаниях они приходят в движение, что и приводит к возникновению нервного импульса. В зависимости от того, какова частота колебания, возбуждаются слуховые клетки, расположенные в разных местах кортиевого органа, что и создает ощущение различной высоты звука.
Раздельное представительство звуков имеется не только на периферическом уровне в кортиевом органе, но и на всех других уровнях слуховой системы, включая и кору больших полушарий. Первичное 41-е поле височной коры принципиально организовано так же, как и первичное зрительное 17-е поле или первичное тактильное 3-е поле: в разных его участках представлены различные участки звуковой тон-шкалы. Тонотопическая организация присуща всей слуховой системе, начиная от кортиева органа улитки, и кончая первичным 41-м полем коры больших полушарий.
При поражении кортиевого органа (вследствие воспалительных или травматических процессов, в частности из-за болезни Миньера) у человека нарушается нормальное восприятие громкости звуков; они или вызывают ощущение боли, или вообще не воспринимаются. В клинике выделяют две основные формы снижения слуха:
♦ первая из них связана с патологическими процессами в среднем ухе (кондуктивная глухота);
♦ вторая — с патологическими процессами во внутреннем ухе (невральная глухота).
Последняя возникает при поражении кортиева органа (а также улиткового нерва). Для нее характерно «явление рекрутмента» — неожиданное появление сильного звукового ощущения (вплоть до болевых ощущений) при плавном нарастании интенсивности звука.
VIII пара черепно-мозговых нервов — очень короткий участок слуховой системы. При поражении VIII нерва (например, при невриномах), который имеет в своем составе и вестибулярные, и слуховые волокна, возникают определенные симптомы, позволяющие однозначно диагностировать поражение этого уровня слуховой системы. К ним относятся различные посторонние звуковые ощущения: шорохи, писк, скрежет и т. п. — и одновременно с ними головокружение. При этом больной хорошо понимает, что реального внешнего источника этих звуков нет, они возникают в его собственном ухе. Иными словами, эти ощущения воспринимаются больным как слуховые обманы. Полная перерезка VIII нерва приводит к полной глухоте на соответствующее ухо.
Следующий уровень слуховой системы — продолговатый мозг (дорсальные и вентральные кохлеарные ядра, где находится второй нейрон слухового пути). В продолговатом мозге происходит первый перекрест путей слуховой системы (переход большинства волокон, несущих слуховую афферентацию, из кохлеарных ядер в ядра верхней оливы и трапециевидного тела своего и противоположного полушария), откуда в составе боковой петли слуховая афферентация попадает в средний мозг, где находятся следующие переключательные ядра слухового пути.
Уровень продолговатого мозга, где находятся несколько ядер, связанных со слуховой рецепцией, очень важен для организации разнообразных безусловных рефлексов, в которых принимают участие звуковые ощущения: рефлекторных движений глаз в ответ на звук, старт-рефлексов в ответ на опасный звук и ряда других безусловных моторных актов, связанных со звуком.
Поражение этого уровня слуховой системы не вызывает нарушений слуха как такового, но ведет к симптомам, связанным с рефлекторной сферой.
Еще одно звено слуховой системы — мозжечок, представляющий своего рода коллектор, собирающий самую различную афферентацию, прежде всего проприоцептивную. Однако в мозжечок поступает и зрительная, и слуховая афферентация. Последняя также имеет большое значение для выполнения основной функции мозжечка — регуляции равновесия. Таким образом, слуховая система, наряду с вестибулярной, участвует и в такой важной функции, как поддержание равновесия.
Важным звеном слуховой системы является средний мозг (нижние бугры четверохолмия). Нижние и верхние бугры четверохолмия тесно взаимодействуют. Здесь на уровне среднего мозга происходят переработка слуховой информации, а также интеграция слуховой и зрительной афферентаций. В области среднего мозга существует частичный перекрест слуховых путей и часть слуховой информации поступает в противоположное полушарие. Именно этот уровень слуховой системы прежде всего участвует в биноуральном слухе, т. е. в способности одновременно оценивать и удаленность, и пространственное расположение источника звука, что делается с помощью сопоставления ощущений, поступающих от левого и правого ушей. Нарушение биноурального слуха является основным симптомом поражения среднего мозга (нижних бугров четверохолмия).
Медиальное или внутреннее коленчатое тело (МКТ), как известно, входит в состав таламической системы, представляющей собой важнейший коллектор различного рода афферентаций, в том числе и слуховой. В разных участках МКТ представлены разные участки тон-шкалы. При поражении МКТ возникают различные нарушения работы слуховой системы, которые, к сожалению, недостаточно хорошо описаны в клинической литературе. Они выражаются прежде всего в снижении способности воспринимать звуки ухом, противоположным очагу поражения, а также в появлении слуховых галлюцинаций.
Следующий уровень — слуховое сияние (пучок Грациоле) — волокна, которые идут из MKT к 41-му первичному полю коры височной области мозга. Слуховое сияние — достаточно большой по протяженности участок слуховой системы, который довольно часто поражается тем или иным патологическим процессом (опухолями, травмой и т. д.); при этом отмечается снижение слуха на противоположное ухо. Имеются указания и на появление в этих случаях (как и при поражении МКТ) слуховых галлюцинаций.
Предполагается, что слуховые галлюцинации (как и зрительные) связаны не с поражением таламического или надталамического уровней слуховой системы, а с раздражением этих областей. В отличие от элементарных звуковых обманов, которые возникают при поражении слухового нерва, в этих случаях появляются сложные слуховые симптомы в виде окликов, бытовых, музыкальных звуков и т. п., т. е. в виде «оформленных», имеющих смысл звуковых образов. Последняя инстанция слухового пути — 41-е первичное поле коры височной области мозга, организованное по топическому принципу, — расположено в извилине Гешеля, в глубине височной коры, и не выходит на поверхность. Во время электрического раздражения первичной слуховой коры больные слышат простые звуки (высокой или низкой частоты), но не слова. Очаг поражения, расположенный в 41-м ноле одного полушария, не приводит к центральной глухоте на соответствующее ухо, так как слуховая афферентация поступает одновременно в оба полушария (преимущественно — в противоположное полушарие). Однако при этом появляются другие симптомы.
[…] корковый уровень слуховой системы связан прежде всего с анализом коротких звуков (меньше 4 мс), что проявляется в виде невозможности восприятия и различения коротких звуков при его поражении; причем этот симптом характерен для поражения как левой, так и правой височных областей.
Все описанные выше нарушения относятся к относительно элементарным сенсорным слуховым расстройствам.
Гностические слуховые расстройства
Гностические слуховые расстройства связаны с поражением ядерной зоны слухового анализатора (куда кроме 41-го поля входят 42-е и 22-е поля). В клинической и нейропсихологической литературе многократно описаны нарушения слуховых функций, возникающие при поражениях ядерной зоны слуховой системы правого и левого полушарий.
При поражении вторичных корковых полей слуховой системы правого полушария (42-го и 22-го) больные (правши) не способны определить значение различных бытовых (предметных) звуков и шумов. Это нарушение носит название слуховая или акустическая агнозия.
В грубых случаях слуховая агнозия выражается в том, что больные не могут определить смысл самых простых бытовых звуков, например скрип дверей, шум шагов, звук льющейся воды и т. п., т. е. всех тех звуков, которые мы привыкли различать без специального обучения. Подобные звуки перестают быть для больных носителями определенного смысла, хотя слух как таковой у них сохранен и они могут различать звуки по высоте, интенсивности, длительности и тембру. В этих случаях наблюдается принципиально такое же нарушение, какое возникает и при зрительной агнозии, когда при полной сохранности остроты и полей зрения нарушается способность понимать увиденное.
Подобные случаи сравнительно редки. Выраженная слуховая агнозия наблюдается при обширном поражении правой височной области, описаны случаи грубой слуховой агнозии при двухстороннем поражении височных областей мозга. По данным некоторых авторов, слуховая агнозия наблюдается при поражении не только субдоминантного, но и доминантного (левого для правшей) полушария. Чаше встречается более стертая форма слуховых нарушений в виде дефектов слуховой памяти. Дефекты слуховой памяти проявляются в специальных экспериментах, показавших, что такие больные, способные различать звуко-высотные отношения, не могут выработать слуховые дифференцировки, т. е. запомнить два (или больше) звуковых эталона. У больных с височными поражениями нарушается также способность к различению звуковых комплексов разной сложности, особенно состоящих из серии последовательных звуков.
При поражении височной области мозга возникает аритмия. Ее симптомы, хорошо изученные А.Р. Лурия и его сотрудниками, состоят в том, что больные не могут правильно оценить и воспроизвести относительно простые ритмические структуры, которые предъявляются им на слух.
В качестве эталонов больным предъявляются наборы звуков (пачки), чередующихся через разные промежутки (или сгруппированных в определенные структуры по 2-3-4-5 звуков в пачке); внутри пачки отдельные удары акцентируются. Больной должен уловить структуру ритмов и «узор» стимулов внутри пачки.
Различение и воспроизведение подобных элементарных ритмических структур для любого здорового человека не представляет никаких сложностей. Больные с поражением височных областей мозга, как правило, не способны оценить количество звуков: они либо переоценивают, либо недооценивают количество ударов, не различая, сколько звуков было в пачке и как они чередовались друг с другом. Эта проба выявляет дефект сенсорной слуховой памяти как таковой, а также дефект различения последовательных комплексных стимулов.
Нужно отметить, что и здоровые испытуемые довольно сильно различаются своими способностями оценки и воспроизведения ритмических структур, особенно если последние предъявляются в быстром темпе.
Неслучайно этот тест используется в музыкальных школах при отборе музыкально одаренных или просто способных к обучению музыке детей.
В клинике применяются сравнительно простые звуковые тесты, иначе можно принять за симптом аритмии немузыкальностъ испытуемых, что является не дефектом, а лишь вариантом нормы.
Один из хорошо описанных в психологической литературе дефектов неречевого слуха называется амузией.
Это нарушение способности узнавать и воспроизводить знакомую или только что услышанную мелодию, а также отличать одну мелодию от другой.
Симптом амузии не совпадает с афазическими расстройствами, что, в частности, было описано А. Р. Лурия и его сотрудниками (1968), показавшими, что у больного может быть резкое расхождение музыкальных и речевых способностей в виде грубой афазии при сохранности музыкального слуха.
Больные с амузией не только не могут узнать мелодию, но и оценивают музыку как болезненное и неприятное переживание. Они рассказывают, что не узнают любимых мелодий, музыка для них потеряла смысл и вызывает приступы головной боли, т. е. стала для них активно неприятной.
Амузия, по-видимому, связана с нарушением не столько звуко-высотного слуха, сколько более сложной способности к музыкальной комбинаторике и к музыкальной грамоте. Больные, обучавшиеся ранее музыке и знавшие музыкальную грамоту, теряют и эти знания.
Важно отметить, что симптом амузии проявляется главным образом при поражении правой височной области, а явления аритмии могут наблюдаться не только при правосторонних, но и при левосторонних височных очагах (у правшей).
Наконец, симптомом поражения правой височной области является, как уже говорилось выше, нарушение интонационной стороны речи. Больные с поражением правой височной области часто не только не различают речевых интонаций, но и не очень выразительны в собственной речи, которая лишена модуляций, интонационного разнообразия, свойственного здоровому человеку. У таких больных часто нарушено пение.
Известны описания больных с поражением правой височной области, которые, правильно повторяя отдельную фразу, не могли ее пропеть, ибо при пении интонационный компонент речи усиливается. Нарушения восприятия интонационных компонентов речи отмечаются и в тех случаях, когда после односторонней электрошоковой терапии угнетаются функции всего правого полушария мозга. В этих случаях человек иногда не может определить на слух даже принадлежность голоса мужчине или женщине.
Если при затормаживании левого полушария вследствие электросудорожной терапии человек становится нечувствительным, невнимательным к речевым звукам (он как бы не слышит того, что ему говорят, хотя полная словесная глухота отсутствует), то при затормаживании правого полушария он слышит речь, но не знает, кто это говорит (мужчина или женщина), и не понимает интонационных характеристик высказывания (вопросительных, утвердительных, восклицательных и т. д.). Эти факты хорошо согласуются с наблюдениями, полученными в клинике. Для больных с поражениями правой височной области мозга также характерно нарушение «эмоционального слуха» — плохое различение интонаций речи, отражающих разные эмоциональные состояния (радость, печаль, гнев и др.).
Физиология человека: Compendium. Учебник для высших учебных заведений / Под ред. Акад РАМН Б.И.Ткаченко и проф. В.Ф.Пятина, СПб. – 1996, 424 с.
Вестибулярная система
Вестибулярная система регулирует равновесие и ориентацию человека в пространстве. Эта система очень чувствительна к линейным ускорениям и к угловым вращениям.
Периферический отдел вестибулярной системы (органы гравитации и равновесия, вестибулярный аппарат) расположен в височной кости и состоит из двух статолитовых (отолитовых) органов — утрикулус (овальный мешочек) и саккулус (круглый мешочек) и трех взаимно перпендикулярных полукружных каналов - горизонтального и вертикальных переднего и заднего. Каждый канал имеет расширение - ампулу. Комплекс утрикулус, саккулус и полукружные каналы называют перепончатым лабиринтом, который заполнен эндолимфой, сообщающейся с эндолимфой улитки, и окружен перилимфой, связанной с перилимфой органа слуха.
Статолитовые органы воспринимают линейные ускорения, а их адекватным стимулом является сила тяжести.
Сенсорный эпителий представлен рецепторными волосковыми, а также опорными клетками. Волоски (цилии) рецепторных клеток обращены в просвет перепончатого лабиринта. Они состоят из подвижных длинных волосков (киноцилий) и коротких менее подвижных, а также многочисленных стереоцилий (порядка 60 на клетке). Вестибулярные афферентные нервные волокна подходят к основанию рецептора и имеют постоянную спонтанную активность (импульсация без воздействия раздражителя).
Волоски рецепторных клеток мешочков погружены в желатинообразную отолитовую мембрану, содержащую мелкие, но тяжелые кристаллы карбоната кальция (отолиты). При прямом положении тела и головы утрикулус находится в горизонтальном, а саккулус - в вертикальном положении. Наклон головы изменяет на некоторый угол положение утрикулуса и саккулуса, смещает мембрану относительно сенсорного эпителия и сгибает цилии. Смещение стереоцилий по направлению к киноцилий усиливает импульсную активность афферентного волокна, а смещение в противоположную сторону от киноцилий снижает частоту спонтанного разряда афферентного волокна. Рецепторы утрикулуса более чувствительны к изменениям положения головы и тела, рецепторы саккулуса — к вибрациям в диапазоне частот до 2000 Гц.
Рецепторы полукружных каналов реагируют на угловые ускорения, т. е. на повороты головы и туловища вокруг вертикальной оси, на наклоны головы вперед, назад, влево и вправо.
Каждый канал имеет расширение (ампулу), содержащее желатинообраз-ный выступ - купулу, выступающий в эндолимфу. В вещество купулы погружены цилии волосковых рецепторных клеток. Купула и эндолимфа имеют одинаковый удельный вес. Поэтому при повороте головы эндолимфа сохраняет прежнее положение, а свободный конец купулы отклоняется в направлении, противоположном повороту, и сгибает цилии волосковых клеток. Это смещение или сгибание цилий является адекватным стимулом для рецепторов полукружных каналов. Рецепторный потенциал волосковых клеток передается к нервным окончаниям (дендритам) биполярных клеток вестибулярного ганглия с помощью медиатора ацетилхолина.
Центральное представительство вестибулярной системы (вестибуло-рецеп-торов). Возбуждение от рецепторов передается по терминальным дендритным волокнам биполярных клеток вестибулярного ганглия к вестибулярным ядрам в продолговатом мозге. От вестибулярных ядер аксоны направляются к мозжечку, ядрам глазодвигательных мышц, вестибулярным ядрам противоположной стороны, к мотонейронам шейного отдела спинного мозга, по вестибулоспинальному тракту - к мотонейронам мышц-разгибателей, а также к ретикулярной формации, гипоталамусу и таламическим ядрам. Функциональное значение этих связей - автоматический контроль (без участия сознания) равновесия тела, поддерживаемого врожденными рефлексами. От зон таламуса (дорзомедиальная часть вентрального постлатерального ядра) сигналы о положении головы и тела поступают к задней постцентральной извилине коры большого мозга. Таламокортикальная проекция обеспечивает сознательный анализ положения тела в пространстве и восприятие перемещений (скорость, ориентация и т. д.). Кроме того, вестибулярная система
представлена также в моторной коре. Афферентация сюда поступает через вестибуло-мозжечково-таламический путь, который переключается в медиальной части вентрального ядра таламуса. Этот опосредованный мозжечковый путь поддерживает тонические реакции, связанные с оценкой позы и со схемой тела.
Чувство равновесия формируется на основании восприятия информации о положении тела и головы в пространстве и схеме тела, которая в текущий момент строится мозгом на основе интеграции афферентной информации от органа равновесия и от проприорецепторов суставов и мышц в сочетании со зрительным контролем ориентировки человека в пространстве. Существенная роль в этом принадлежит врожденным рефлексам.
Вестибулярные раздражения вызывают статические и статокинетические рефлексы. Статические рефлексы поддерживают равновесие при положениях тела стоя и разных углах наклона. Они осуществляются с рецепторов отолитовых органов (утрикулус и саккулус). Статокинетические рефлексы регулируют тонус мышц во время движений (например, поворот тела при свободном падении) и обеспечиваются как статолитовыми органами, так и рецепторами полукружных каналов.
Среди статокинетических рефлексов особое значение имеет вестибулярный нистагм (вестибуло-окуломоторная реакция) или серия ритмических движений глаз в сторону, противоположную вращению тела. Нистагм глаз поддерживает восприятие стабильной картины внешнего мира.
Вестибулярная система играет важную функциональную роль в регуляции и контроле моторных вестибулоспинальных и вестибуловисцеральных реакций. Вестибулоспинальные реакции перераспределяют мышечный тонус и поддерживают равновесие через вестибуло- рубро- и ретикулоспинальные тракты на сегментарном уровне. Вестибулоспинальные реакции (быстрые, срочные) находятся под контролем мозжечка. Вестибуловисцеральные реакции выполняют структуры продолговатого мозга, ствола и среднего мозга. Они проявляются в изменении работы желудочно-кишечного тракта (тошнота, рвота), сердечно-сосудистой системы (дизритмшг), возникающих при нагрузках на вестибулярную систему (болезнь движения, морская болезнь).
Смирнов В.М., Будылина С.М. Физиология сенсорных систем и высшая нервная деятельность: Учеб. пособие для студ. высш. учеб. заведений. – М.: Академия, 2003. – с. 63 – 67
Вестибулярный анализатор
Вестибулярный анализатор обеспечивает так называемое акселерационное чувство, т. е, ощущение, возникающее при прямолинейном и вращательном ускорении движения тела, а также при изменениях положения головы. Вестибулярному анализатору принадлежит ведущая роль в пространственной ориентации человека, сохранении его позы.
Структурно-функциональная характеристика. Периферический (рецепторный) отдел вестибулярного анализатора представлен во-шоковыми клетками вестибулярного органа, расположенного, как и улитка, в лабиринте пирамиды височной кости. Вестибулярный орган (орган равновесия, орган гравитации) состоит из трех полукружных каналов и преддверия.
Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях: верхний — во фронтальной, задний — в сагиттальной и наружный — в горизонтальной. Преддверие состоит из двух мешочков: круглого (саккулюс), расположенного ближе улитке, и овального (утрикулюс), расположенного ближе к полукружным каналам. Полукружные каналы своими устьями открываются в преддверие и сообщаются с ним пятью отверстиями (колено двух каналов, а именно верхнего и заднего, соединены вместе). Один конец каждого канала имеет расширение, которое называется ампулой. Все эти структуры состоят из тонких перепонок и образуют перепончатый лабиринт, внутри которого находится эндолимфа. Вокруг перепончатого лабиринта и между ним и его костным футляром имеется перилимфа, которая переходит в перилимфу органа слуха. В каждом мешочке преддверия имеются небольшие возвышения, называемые пятнами, а в ампулах полукружных каналов — гребешками. Они состоят из нейроэпителиальных клеток, имеющих на свободной поверхности волоски (реснички), которые разделяются на две группы: тонкие, их много, — стереоцилии и один более толстый и длинный на периферии пучка — киноцилии.

Рисунок: вестибулярный орган
Волосковые клетки представляют собой рецепторы вестибулярного анализатора и являются вторичными. Рецепторные клетки преддверия покрыты желеобразной массой, которая состоит в основном из мукополисахаридов, благодаря содержанию значительного количества кристаллов карбоната кальция получила название отолитовой мембраны. В ампулах полукружных каналов желеобразная масса не содержит солей Ткальция и называется листовидной мембраной (купулой). Волоски рецепторных клеток пронизывают эти мембраны. Возбуждение колосковых клеток происходит вследствие скольжения мембраны по волоскам, изгибания волосков (стереоцилии) в сторону киноцилии. При этом возникает рецепторный потенциал волосковых клеток и выделяется медиатор ацетилхолин, который стимулирует синаптические окончания волокон вестибулярного нерва. Этот эффект проявляется в усилении постоянной спонтанной активности вестибулярного нерва. Если же смещение стереоцилии направлено в противоположную от киноцилии сторону, то спонтанная активность вестибулярного нерва снижается.
Для волосковых клеток преддверия адекватными раздражителями являются ускорение или замедление прямолинейного движения тела, а также наклоны головы. Под действием ускорения отолитовая мембрана скользит по волосковым клеткам, а при изменении положения головы меняет свое положение по отношению к ним. Это вызывает отклонение ресничек и возникновение возбуждения в рецепторных волосковых клетках. Порог различения ускорения равен 2 – 20 см/с. Порог различения наклона головы в сторону составляет около 1°, а вперед и назад – около 2°. При сопутствующих раздражениях (вибрация, качка, тряска) происходит снижение чувствительности вестибулярного аппарата. Так, вибрации, имеющие место в самолетах, повышают порог различения наклона головы вперед и назад до 5°, при наклонах в стороны – до 10°.
Для волосковых клеток полукружных каналов адекватным раздражителем является ускорение или замедление вращательного движения в какой-либо плоскости. Поскольку полукружные каналы заполнены эндолимфой, имеющей такую же плотность, как и купула ампул, линейные ускорения не оказывают влияния на положение ресничек и купулы. При поворотах головы или вращении тела, т.е. при появлении углового ускорения, эндолимфа в них в силу своей инерции в первый момент остается неподвижной или потом движется, но с иной скоростью, нежели полукружные каналы. Это вызывает сгибание ресничек рецепторов в купуле и возбуждение их. В зависимости от характера вращательного ускорения или замедления происходит неодинаковое раздражение рецепторов различных полукружных каналов. По картине импульсов, приходящих в центральные структуры вестибулярного анализатора из полукружных каналов с каждой стороны, мозг получает информацию о характере вращательного движения. Рецепторы полукружных каналов дают возможность различать угловое ускорение.
Проводниковый отдел. К рецепторам подходят периферические волокна биполярных нейронов вестибулярного ганглия, расположенного во внутреннем слуховом проходе (первый нейрон). Аксоны этих нейронов в составе вестибулярного нерва направляются к вестибулярным ядрам продолговатого мозга (второй нейрон). Вестибулярные ядра продолговатого мозга (верхнее — ядро Бехтерева, медиальное — ядро Швальбе, латеральное — ядро Дейтерса и нижнее — ядро Роллера) получают дополнительную информацию по афферентным нейронам от проприорецепторов мышц или от суставных сочленений шейного отдела позвоночника. Эти ядра вестибулярного анализатора тесно связаны с различными отделами центральной нервной системы. Благодаря этому обеспечиваются контроль и управление эффекторными реакциями соматического, вегетативного и сенсорного характера. Третий нейрон расположен в ядрах зрительного бугра, откуда возбуждение направляется в кору полушарий.
Центральный отдел вестибулярного анализатора локализуется в височной области коры большого мозга, несколько кпереди от слуховой проекционной зоны (21 — 22 поля по Бродману, четвертый нейрон).
Функциональные связи вестибулярного анализатора. При возбуждении вестибулярного анализатора возникают соматические реакции, которые осуществляются благодаря вестибулоспинальным связям при участии вестибулоретикулярных и вестибулоруброспинальных трактов. При этом происходят перераспределение тонуса скелетной мускулатуры и рефлекторные реакции, необходимые для сохранения равновесия тела в пространстве. Рефлексы, обеспечивающие данную функцию, подразделяются на две группы — статические и статокинетические.
Один из статокинетических рефлексов — вестибулярный нистагм (головы или глаз) — имеет большое клиническое значение. Нистагм возникает в условиях быстрого перемещения тела или его вращения. Так, глазной нистагм проявляется сначала в ритмическом медленном движении глаз в сторону, противоположную вращению, а затем — быстром движении глаз (скачком) в обратном направлении. Реакции такого типа обеспечивают возможность обзора пространства в условиях перемещения тела. Важным моментом является связь вестибулярного аппарата с мозжечком, благодаря чему осуществляется тонкая регуляция моторных вестибулярных рефлексов. При нарушениях функции мозжечка эти рефлексы утрачивают тормозной компонент, что проявляется в возникновении таких симптомов, как, например, спонтанно возникающий нистагм, утрата равновесия, избыточная амплитуда движений. Эти симптомы являются частью синдрома мозжечковой атаксии. Благодаря связям вестибулярных ядер с вегетативной нервной системой проявляются вестибуловегетативные реакции сердечно-сосудистой системы, желудочно-кишечного тракта и других органов. Они могут проявляться в изменениях сердечного ритма, тонуса сосудов, артериального давления, усилении моторики желудка и кишечника, повышении саливации, тошноте, рвоте и т.д. В условиях невесомости (в космосе) возникает такой тип афферентной импульсации с вестибулярного аппарата, который никогда не встречается на Земле. Однако привыкание к условиям невесомости во время космических полетов происходит быстро. При этом следует учитывать, что космонавты проходят напряженный курс тренировки, чем и объясняется их малая подверженность влиянию условий невесомости.
Психофизиология. Под ред. Ю.И. Александрова. – СПб.: Питер, 2003
Вестибулярная система
Вестибулярная система играет важную роль в пространственной ориентации человека. Она получает, передает и анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения, а также при изменении положения головы в пространстве. При равномерном движении или в условиях покоя рецепторы вестибулярной системы не возбуждаются. Импульсы от вестибулорецепторов вызывают перераспределение тонуса скелетной мускулатуры, что обеспечивает сохранение равновесия тела.
Строение и функции рецепторного вестибулярного аппарата
Периферическим отделом вестибулярной системы является вестибулярный аппарат, расположенный в пирамиде височной кости. Он состоит из преддверия и трех полукружных каналов. Полукружные каналы располагаются в трех взаимно перпендикулярных плоскостях. Один из концов каждого канала расширен (ампула). Вестибулярный аппарат включает в себя также два мешочка. В них на возвышениях находится отолитовый аппарат: скопления рецепторных клеток. Выступающая в полость мешочка рецепторная клетка оканчивается длинным подвижным волоском и 60-80 склеенными неподвижными волосками. Они пронизывают желеобразную мембрану, содержащую кристаллики карбоната кальция — отолиты. Волосковые клетки возбуждаются при скольжении отолитовой мембраны по волоскам, т. е. при их сгибании.
В перепончатых полукружных каналах, заполненных эндолимфой, рецепторные волосковые клетки сконцентрированы в ампулах. Во время угловых ускорений эндолимфа приходит в движение, волоски сгибаются и волосковые клетки возбуждаются. При противоположно направленном движении они тормозятся. Это связано с тем, что отклонение волоска в одну сторону приводит к открыванию каналов и деполяризации волосковой клетки, а отклонение в противоположном направлении закрывает каналы и гиперполяризует рецептор. В волосковых клетках преддверия и ампулы при их сгибании генерируется рецепторный потенциал, который усиливает выделение ацетилхолина и через синапсы активирует окончания волокон вестибулярного нерва.
Волокна вестибулярного нерва (отростки биполярных нейронов) направляются в продолговатый мозг. Импульсы, приходящие по этим волокнам, активируют нейроны бульбарного вестибулярного комплекса (ядра: преддверное верхнее Бехтерева, преддверное латеральное Дейтерса, Швальбе и др.). Отсюда сигналы направляются во многие отделы ЦНС: спинной мозг, мозжечок, глазодвигательные ядра, кору мозга, ретикулярную формацию и вегетативные ганглии.
Рефлексы, связанные с вестибулярной стимуляцией
Нейроны вестибулярных ядер обеспечивают контроль над различными двигательными реакциями и управление ими. Вестибуло-спинальные влияния изменяют импульсацию нейронов сегментарных уровней спинного мозга. Так осуществляется динамическое перераспределение тонуса скелетной мускулатуры и включаются рефлекторные реакции, необходимые для сохранения равновесия. В вестибуловегетативные реакции вовлекаются сердечно-сосудистая система, желудочно-кишечный тракт и другие внутренние органы. При сильных и длительных нагрузках на вестибулярный аппарат возникает болезнь движения (например, морская болезнь). Вестибуло-глазодвигательные рефлексы (глазной нистагм) состоят в медленном ритмическом движении глаз в противоположную вращению сторону, сменяющемся их скачком обратно. Возникновение и характеристики вращательного глазного нистагма — важные показатели состояния вестибулярной системы и широко используются в эксперименте и клинике.
Основные афферентные пути и проекции вестибулярных сигналов
Два основных пути поступления вестибулярных сигналов в кору мозга обезьян следующие: прямой — через вентральное постлатеральное ядро и непрямой — через вентролатеральное ядро. В коре основные афферентные проекции вестибулярного аппарата локализованы в задней части постцентральной извилины. В моторной коре кпереди и книзу от центральной борозды обнаружена вторая вестибулярная зона. Локализация вестибулярной зоны в коре мозга человека окончательно не выяснена.
Функции вестибулярной системы
Вестибулярная система помогает ориентироваться в пространстве при активном и пассивном движении. При пассивном движении лабиринтный аппарат с помощью корковых отделов системы анализирует и запоминает направление движения и повороты. Следует подчеркнуть, что в нормальных условиях пространственная ориентировка обеспечивается совместной деятельностью зрительной и вестибулярной систем.
Чувствительность вестибулярной системы здорового человека очень высока: отолитовый аппарат позволяет воспринять ускорение прямолинейного движения, равное всего 2 см/с2. Порог различения наклона головы в сторону — всего около 1 углового градуса, а вперед и назад — 1,5-2 угловых градуса.