

Раздел «Анатомия» портала http://medicinform.Net
Физиология конечного мозга
Конечный мозг, или полушария большого мозга, достигшие своего наивысшего развития у человека, справедливо считается самым сложным и самым удивительным созданием природы.
Функции этого отдела центральной нервной системы настолько отличаются от функций ствола и спинного мозга, что они выделяются в особую главу физиологии, называемую высшей нервной деятельностью. Этот термин введен И.П. Павловым. Деятельность нервной системы, направленную на объединение и регуляцию всех органов и систем организма, И.П. Павлов назвал низшей нервной деятельностью. Под высшей нервной деятельностью он понимал поведение, деятельность, направленную на приспособление организма к изменяющимся условиям внешней среды, на уравновешивание с окружающей средой. В поведении животного, в его взаимоотношениях с окружающей средой ведущую роль играет конечный мозг, орган сознания, памяти, а у человека - орган умственной деятельности, мышления.
[…]
Для изучения локализации (места нахождения) функций в коре полушарий большого мозга, или, иными словами, значения отдельных зон коры, применяют различные методы: частичное удаление коры, электрическое и химическое раздражение, запись биотоков мозга и метод условных рефлексов.
Метод раздражения позволил установить в коре следующие зоны: двигательные (моторные), чувствительные (сенсорные) и немые, которые теперь называют ассоциативными.
Двигательные (моторные) зоны коры.
Движения возникают при раздражении коры в области предцентральной извилины. Электрическое раздражение верхней части извилин вызывает движение мышц ног и туловища, средней - рук, нижней - мышц лица.
Величина корковой двигательной зоны пропорциональна не массе мышц, а точности движений. Особенно велика зона, управляющая движениями кисти руки, языком, мимической мускулатурой лица. В V слое коры двигательных зон обнаружены гигантские пирамидные клетки (пирамиды Беца), отростки которых спускаются к двигательным нейронам среднего, продолговатого и спинного мозга, иннервирующим скелетную мускулатуру.
Путь от коры к двигательным нейронам носит название пирамидного пути. Это путь произвольных движений. После повреждения моторной зоны произвольные движения не могут осуществляться.
Раздражение моторной зоны сопровождается движениями на противоположной половине тела, что объясняется перекрестом пирамидных путей на их пути к двигательным нейронам, иннервирующим мышцы.
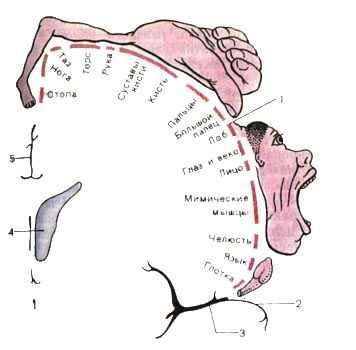
Рисунок: двигательный гомункулус. Показаны проекции частей тела человека на область коркового конца двигательного анализатора.
Сенсорные зоны коры.
Экстирпация (искоренение) различных участков коры у животных позволила в общих чертах установить локализации сенсорных (чувствительных) функций. Затылочные доли оказались связанными со зрением, височные - со слухом.
Зона коры, куда проецируется данный вид чувствительности, называется первичной проекционной зоной.
Кожная чувствительность человека, чувства прикосновения, давления, холода и тепла проецируются в постцентральную извилину. В верхней ее части находится проекция кожной чувствительности ног и туловища, ниже - рук и совсем внизу - головы.
Абсолютная величина проекционных зон отдельных участков кожи неодинакова. Так, например, проекция кожи кисти рук занимает в коре большую площадь, чем проекция поверхности туловища.
Величина корковой проекции пропорциональна значению данной рецептивной поверхности в поведении. Интересно, что у свиньи особенно велика проекция в кору пятачка.
Суставно-мышечная, проприоцептивная, чувствительность проецируется в постцентральную и предцентральную извилины.
Зрительная зона коры находится в затылочной доле. При раздражении ее возникают зрительные ощущения - вспышки света; удаление ее приводит к слепоте. Удаление зрительной зоны на одной половине мозга вызывает слепоту на одной половине каждого глаза, так как каждый зрительный нерв делится в области основания мозга на две половины (образует неполный перекрест), одна из них идет к своей половине мозга, а другая – к противоположной.
При повреждении наружной поверхности затылочной доли не проекционной, а ассоциативной зрительной зоны зрение сохраняется, но наступает расстройство узнавания (зрительная агнозия). Больной, будучи грамотным, не может прочесть написанное, узнает знакомого человека после того, как тот заговорит. Способность видеть – это врожденное свойство, но способность узнавать предметы вырабатывается в течение жизни. Бывают случаи, когда от рождения слепому возвращают зрение уже в старшем возрасте. Он еще долгое время продолжает ориентироваться в окружающем мире на ощупь. Проходит немало времени, пока он научится узнавать предметы с помощью зрения.
 Рисунок:
чувствительный гомункулус. Показаны
проекции частей тела человека на область
коркового конца анализатора.
Рисунок:
чувствительный гомункулус. Показаны
проекции частей тела человека на область
коркового конца анализатора.
Функция слуха обеспечивается точными долями больших полушарий. Раздражение их вызывает простые слуховые ощущения.
Удаление обеих слуховых зон вызывает глухоту, а одностороннее удаление понижает остроту слуха. При повреждении участков коры слуховой зоны может наступить слуховая агнозия: человек слышит, но перестает понимать значение слов. Родной язык становится ему так же непонятен, как и чужой, иностранный, ему незнакомый. Заболевание носит название слуховой агнозии.
Обонятельная зона коры находится на основании мозга, в области парагиппокампальной извилины.
Проекция вкусового анализатора, по-видимому, находится в нижней части постцентральной извилины, куда проецируется чувствительность полости рта и языка.
Лимбическая система.
В конечном мозге располагаются образования (поясная извилина, гиппокамп, миндалевидное тело, область перегородки), составляющие лимбическую систему. Они участвуют в поддержании постоянства внутренней среды организма, регуляции вегетативных функций и формировании эмоций и мотиваций. Эту систему иначе называют «висцеральным мозгом», так как эта часть конечного мозга может рассматриваться как корковое представительство интерорецепторов. Сюда поступает информация от внутренних органов. При раздражении желудка, мочевого пузыря в лимбической коре возникают вызванные потенциалы.
Электрическое раздражение различных областей лимбической системы вызывает изменения вегетативных функций: кровяного давления, дыхания, движений пищеварительного тракта, тонуса матки и мочевого пузыря.
Разрушение отдельных частей лимбической системы приводит к нарушению поведения: животные могут становиться более спокойными или, напротив, агрессивными, легко дающими реакции ярости, изменяется половое поведение. Лимбическая система имеет широкие связи со всеми областями головного мозга, ретикулярной формацией и гипоталамусом. Она обеспечивает высший корковый контроль всех вегетативных функций (сердечно-сосудистой, дыхательной, пищеварительной, обмена веществ и энергии.

Рисунок: образования головного мозга, относящиеся к лимбической системе (круг Папеца).
1 - обонятельная луковица; 2 - обонятельный путь; 3 - обонятельный треугольник; 4 - поясная извилина; 5 - серые включения; 6 - свод; 7 - перешеек поясной извилины; 8 - концевая полоска; 9 - гиппокампальная извилина; 11 - гиппокамп; 12 - сосцевидное тело; 13 - миндалевидное тело; 14 - крючок.
Ассоциативные зоны коры.
Проекционные зоны коры занимают в мозге человека небольшую долю всей поверхности коры. Остальная поверхность занята так называемыми ассоциативными зонами. Нейроны этих областей не связаны ни с органами чувств, ни с мышцами, они осуществляют связь между различными областями коры, интегрируя объединяя все притекающие в кору импульсы в целостные акты научения (чтение, речь, письмо), логического мышления, памяти и обеспечивая возможность целесообразной реакции поведения.
При нарушениях ассоциативных зон появляются агнозии - неспособность узнавания и апраксии - неспособность производить заученные движения. Например, стереоагнозия выражается в том, что человек не может найти на ощупь у себя в кармане ни ключа, ни коробки спичек, хотя зрительно он их сейчас же узнает. Выше приводились примеры зрительной агнозии - неспособность прочесть написанное и слуховой - непонимание значения слов.
При нарушении ассоциативных зон коры может наступить афазия - потеря речи. Афазия может быть моторной и сенсорной. Моторная афазия возникает при поражении задней трети нижней лобной извилины слева, так называемого центра Брока (этот центр находится только в левом полушарии). Больной понимает речь, но сам говорить не может. При сенсорной афазии, поражении центра Вернике в задней части верхней височной извилины, больной речи не понимает.
При аграфии человек разучивается писать, при апраксии - производить заученные движения: зажечь спичку, застегнуть пуговицу, пропеть мелодию и др.
Изучение локализации функции методом условных рефлексов на живом здоровом животном позволило И.П. Павлову обнаружить факты, на основе которых им была построена теория динамической локализации функций в коре, затем блестяще подтвержденная при помощи микроэлектродного исследования нейронов. У собак вырабатывали условные рефлексы, например на зрительные раздражения - свет, различные фигуры - круг, треугольник, а затем удаляли всю затылочную, зрительную, зону коры. После этого условные рефлексы исчезали, но проходило время, и нарушенная функция частично восстанавливалась. Это явление компенсации, или восстановления, функции И.П. Павлов объяснил, высказав мысль о существовании ядра анализатора, расположенного в определенной зоне коры, и рассеянных клеток, разбросанных по всей коре, в зонах других анализаторов. За счет этих сохранившихся рассеянных элементов происходит восстановление утраченной функции. Собака может отличать свет от тьмы, но тонкий анализ, установление различий между кругом и треугольником, ей недоступен, он свойствен только ядру анализатора.
Микроэлектродное отведение потенциалов от отдельных нейронов коры подтвердило наличие рассеянных элементов. Так, в двигательной зоне коры обнаружили клетки, дающие разряд импульсов на зрительные, слуховые, кожные раздражения, а в зрительной зоне коры выявлены нейроны, отвечающие электрическими разрядами на осязательные, звуковые, вестибулярные и обонятельные раздражители. Кроме того, были найдены нейроны, которые отвечают не только на «свой» раздражитель, как теперь говорят, раздражитель своей модальности, своего качества, но и на один – два чужих. Их назвали полисенсорными нейронами.
Динамическая локализация, т. е. способность одних зон замещаться другими, обеспечивает коре высокую надежность.
Общий курс физиологии человека и животных в 2 кн. Кн. 1. Физиология нервной, мышечной и сенсорной систем: Учеб. для биол. и медиц. спец. вузов/ А.Д. Ноздрачев, И.А. Баранникова, А.С. Батуев и др.; Под ред. А.Д. Ноздрачева. – М.: Высш. шк., 1991. – 512 с.
Морфофункциональная организация коры больших полушарий
Полушария конечного мозга состоят из белого вещества, покрытого снаружи серым, или корой, толщина которой в различных отделах больших полушарий колеблется от 1,3 до 4,5 мм. Кора представляет собой филогенетически наиболее молодой и вместе с тем сложный отдел мозга, предназначенный для обработки сенсорной информации, формирования двигательных команд и интеграции сложных форм поведения. Бурный рост неокортекса у высших позвоночных в ограниченном объеме черепа сопровождается образованием многочисленных складок, увеличивающих общую площадь коры, которая у человека составляет 2200 см.
На этом пространстве сконцентрировано 10 – 10 нейронов и еще большее количество глиальных клеток, выполняющих ионорегулирующую и трофическую функции. Образующие кору нейроны по своей геометрии и функции подразделяются на несколько групп. Одну группу составляют варьирующие по размеру пирамидные клетки. Они ориентированы вертикально по отношению к поверхности коры и имеют тело треугольной формы (рис. 3.16). От тела пирамидной клетки вверх отходит длинный Т-образно ветвящийся апикальный дендрит, а вниз от основания нейрона – аксон, который либо покидает кору в составе нисходящих путей, либо направляется к другим зонам коры. Апикальные и более короткие базальные дендриты пирамидных клеток густо усеяны мелкими (до 3 мкм) выростами — шипиками, каждый из которых представляет собой область синаптического контакта.
Другая группа корковых нейронов представлена более мелкими звездчатыми клетками. Эти клетки имеют короткие сильно ветвящиеся дендриты и аксоны, формирующие внутрикорковые связи. Дендриты звездчатых клеток также могут быть снабжены шипиками, которые в процессе онтогенетического развития у человека появляются только к моменту рождения.
Наконец, третья группа корковых нейронов включает в себя вере-теновидные клетки, имеющие длинный аксон, который ориентирован в горизонтальном или вертикальном направлении. В связи с тем, что тела и отростки описанных выше нейронов имеют упорядоченное расположение, кора построена по экранному принципу и у млекопитающих в типичном случае состоит из шести горизонтальных слоев.
Самый наружный молекулярный слой слагается из густого сплетения нервных волокон, лежащих параллельно поверхности корковых извилин. Основную массу этих волокон составляют ветвящиеся апикальные дендриты пирамидных клеток нижележащих слоев. Сюда же в наружный слой приходят афферентные таламокортикальные волокна от неспецифических ядер таламуса, регулирующих уровень возбудимости корковых нейронов.
Второй слой — наружный зернистый — состоит из большого количества мелких звездчатых клеток, которые в вентральной части слоя дополняются малыми пирамидными клетками.
Третий слой — наружным пирамидный — формируется из пирамидных клеток средней величины. Функционально второй и третий слои коры объединяют нейроны, отростки которых обеспечивают кортико-кортикальные ассоциативные связи.
Четвертый слой — внутренний зернистый — содержит множество звездчатых клеток (клеток-зерен), обусловливающих его гранулярную структуру. В этом слое преимущественно оканчиваются афферентные таламокортикальные волокна, идущие от специфических (проекционных) ядер таламуса.
Пятый слой — внутренний пирамидный — образован крупными пирамидными клетками. Наиболее крупные пирамидные нейроны — гигантские клетки Беца — встречаются в прецентральной извилине, занятой моторной зоной коры больших полушарий. Аксоны этих эфферентных корковых нейронов формируют кортикоспинальный (пирамидный) и кортикобульбарные тракты, участвующие в координации целенаправленных двигательных актов и позы.
И наконец, шестой слой — полиморфный, или слой веретеновидных клеток, переходящий непосредственно в белое вещество больших полушарий. Этот слой содержит тела нейронов, чьи отростки формируют кортикоталамические пути.
Такой шестислойный план строения характерен для всего неокортекса. Однако выраженность отдельных слоев в различных областях коры не одинакова. Учитывая эту особенность, К. Бродман по гистологическим признакам, в частности по плотности расположения и форме нейронов, разделил всю кору на 50 цитоархитектонических полей. Позднее были разработаны функциональные принципы классификации различных зон коры. При этом оказалось, что зоны, выделенные на основании их функциональных и нейрохимических особенностей, в известной степени соответствуют цитоархитектоническому разделению коры на поля.
Так, например, при сравнении наиболее изученных сенсорных и моторных зон коры оказалось, что в первых наружный пирамидный слой (3) выражен слабо и доминируют зернистые слои (2, 4), где оканчиваются сенсорные афференты (гранулярная кора). И напротив, в моторных зонах коры зернистые слои развиты плохо (агранулярная кора), а пирамидные слои превалируют.
Таким образом, функциональная специализация накладывает определенный отпечаток на структуру сенсорных и моторных зон коры, и выделение этих областей по различным системам классификации не случайно.
Проекционные зоны коры
Использование методов удаления и раздражения отдельных зон коры позволило идентифицировать ее моторные (мотосенсорные) области. Важнейшая двигательная область коры локализована в прецентральной извилине (поля 4 и 6, по Бродману), лежащей кпереди от центральной (роландовой) борозды, которая отделяет лобную долю от теменной. Слабое электрическое раздражение конкретных точек прецентральной извилины вызывает сокращение определенных групп мышц на противоположной стороне тела.
В 30-х гг. нашего столетия У. Пенфилд, базируясь на результатах, полученных во время нейрохирургических операций, установил наличие правильной пространственной проекции соматических мышц различных отделов тела на двигательную область коры. Обнаружив соматотопическую организацию двигательной коры, У. Пенфилд отметил, что зоны, управляющие движениями кистей рук и мимической мускулатурой лица, являются наиболее обширными и характеризуются самым низким порогом. В дальнейшем на медиальной поверхности полушария рядом с этой первичной моторной областью была обнаружена еще одна вторичная двигательная область и, таким образом, было показано множественное представительство периферических образований в нескольких двигательных зонах коры. В связи с тем что эти области помимо моторного выхода из коры имеют самостоятельные сенсорные входы от кожных и мышечных рецепторов, они были названы первичной и вторичной мотосенсорной корой (Mel и McII).
Последующие экспериментальные исследования показали, что в лежащей позади роландовой борозды постцентральной извилине (поля 1, 2, 3) находится первая соматосенсорная (сенсомоторная) область, куда через специфические ядра таламуса приходят афферентные проекции от рецепторов кожи и двигательного аппарата. Так же как и в двигательной коре, эти проекции имеют соматотопическую организацию, причем близкие друг от друга точки на поверхности тела имеют представительство в соседних участках постцентральной извилины. Удаление определенных зон постцентральной извилины приводит к утрате дискриминационной чувствительности в тех или иных частях тела. Электрическая стимуляция постцентральной извилины у больных при нейрохирургических операциях (под местным наркозом) вызывает чувство онемения или покалывания в определенных местах на поверхности тела.
Кроме первой соматосенсорной области у хищников и приматов обнаружена вторая соматосенсорная область, локализованная вентральнее в районе сильвиевой борозды, которая отделяет височную долю от теменной и лобной. Здесь также имеется соматотопическая проекция поверхности тела, однако менее четкая, чем в первой соматосенсорной области. Оказалось, что первая и вторая соматосенсорные области кроме афферентных входов содержат моторные выходы и, следовательно, их правильнее называть первичной и вторичной сенсомоторными зонами (См1 и CmII).
Следовательно, можно судить о наличии в коре четырех чувствительных областей (CmI, CmII, Mel и McII), расположенных здесь в порядке убывания их значимости. Равным образом в коре существует четыре двигательных области (Mel, McII, CmI, CmII), причем ведущая роль в этом ряду принадлежит первичной мотосенсорной коре.
Из числа других проекционных зон коры следует отметить первичную зрительную область (поле 17), локализованную в затылочной доле. В проекциях рецепторов сетчатки на поле 17, так же как и в других сенсорных системах, обнаруживается точный топографический порядок, причем в каждое полушарие проецируются одноименные половины сетчаток (левые половины — в правое полушарие, а правые — в левое). Происходящее при этом в каждом полушарии совмещение зрительных полей обоих глаз лежит в основе бинокулярного зрения. Повреждение ограниченной области зрительной коры может привести к частичной слепоте, проявляющейся в нарушении небольшой части поля зрения. Электрическое раздражение зрительной коры может вызвать зрительные ощущения.
Рядом с первичной зрительной областью расположена вторичная, занимающая поля 18 и 19. Эти поля имеют отношение к таким функциям, как зрительное внимание и управление движениями глаз.
В настоящее время хорошо изучена первичная проекционная зона слухового анализатора, которая локализована в верхнем крае височной доли (поля 41 и 42). Для этой области, так же как и для других проекционных зон, описано тонотопическое представительство кортиева органа улитки, состоящее в том, что различные участки кортиева органа проецируются в определенные локусы слуховой коры. Повреждение полей 41 и 42 затрудняет восприятие речи, пространственную локализацию источника звука и распознавание временных характеристик звука.
В пределах каждой доли коры больших полушарий рядом с проекционными зонами расположены поля, которые не связаны с выполнением какой-либо специфической сенсорной или моторной функции. Такие поля составляют ассоциативную кору, для нейронов которой свойственно отвечать на раздражение различных модальностей и таким образом участвовать в интеграции сенсорной информации и в обеспечении связей между чувствительными и двигательными зонами коры. Эти механизмы являются физиологической основой высших психических функций.
В состав ассоциативный коры входит ряд областей теменной, височной и лобной долей. К сожалению, границы этих областей и кортико-кортикальные связи между ними идентифицированы не полностью, а большинство физиологических данных базируется на симптомах поражения ассоциативных областей у животных и на данных клинических наблюдений.
Вместе с тем в настоящее время установлено, что теменные ассоциативные поля, которые в процессе эволюции надстраиваются над зрительной проекционной зоной, участвуют в оценке биологически значимой информации и в восприятии пространственных отношений окружающего мира.
Лобные доли (поля 9 – 14), имеющие обширные двусторонние связи с лимбической системой мозга, контролируют оценку мотивации поведения и программирование сложных поведенческих актов. Установлено участие лобных долей в управлении движениями. Так, например, поражение лобных долей вызывает у больных тенденцию к повторению двигательных актов без видимого соответствия с внешними обстоятельствами.
Эксперименты на обезьянах показали, что повреждение лобных долей нарушает запоминание локализации предметов, причем этот процесс связан с дофаминэргическими структурами, которыми богаты лобные доли.
Очевидно, лобные доли наряду с теменными участвуют в интеграции сведений о пространстве и времени.
Физиология ч-ка: Compendium. Учебник для высших учебных заведений / Под ред. Акад РАМН Б.И.Ткаченко и проф. В.Ф.Пятина, СПб. – 1996. – 424 с.
Базальные ганглии
Базальные ганглии головного мозга (стриарные тела) - это три парных образования: неостриатум (хвостатое ядро и скорлупа), палеостриатум (бледный шар) и ограда.
Функции неостриатума. Неостриатум получает афференты от экстрапирамидной и других областей коры. Аксоны нейронов хвостового ядра и скорлупы идут к бледному шару, отсюда — к таламусу и только от него — к сенсорным полям, образуя замкнутый круг: неостриатум – палеостриатум — таламус — кора — неостриатум. Вне этого круга неостриатум связан с черной субстанцией, красным ядром, люисовым телом, вестибулярными ядрами, мозжечком, γ-клетками спинного мозга. На нейронах хвостатого ядра конвергирует информация различных сенсорных систем, т. е. неостриатум является подкорковым интегративным и ассоциативным центром. Обилие и характер связей неостриатума обеспечивают его участие в организации и регуляции движений, работы вегетативных органов.
В норме неостриатум преимущественно тормозит нейроны палеостриату-ма. Присутствие дофамина, который синтезируется в черной субстанции и транспортируется к синапсам нейронов хвостатого ядра, подавляет фоновую активность большинства нейронов хвостатого ядра и частично снимает его тормозящее действие на активность бледного шара. Дофамин регулирует растормаживающий механизм взаимодействия между нео- и палеостриату-мом. При недостатке дофамина в неостриатуме нейроны бледного шара растормаживаются, активизируют спиностволовые системы, что приводит к двигательным нарушениям в виде ригидности мышц,
Неостриатум и палеостриатум принимают участие в таких интегративных процессах как условно-рефлекторная деятельность, двигательная активность, при этом специфичным для активности хвостатого ядра является торможение активности коры, подкорки, безусловного и условно-рефлекторного поведения, прекращение ориентировочной, эмоциональной, двигательной, пищевой активности человека, торможение выработки рефлексов, прекращение восприятия болевых, зрительных, слуховых и других видов сенсорной стимуляции.
Одной из функций хвостатого ядра является регуляция перехода от одного вида движения к другому, т. е. прекращение одного движения и обеспечение нового путем создания необходимой позы, условий для изолированных движений.
Стриопаялидарные ядра регулируют тонус мускулатуры. При повреждении ядер возникают гиперкинезы: непроизвольные мимические реакции, тремор, аттетоз, торсионный спазм, хорея (подергивания конечностей и туловища), длительная гиперактивность в форме бесцельного перемещения с места на место.
Скорлупа, помимо организации двигательной активности, участвует в организации пищевого поведения и в его интеграции с функциями дыхания, слюноотделения и т. д.
Функции палеостриатума (бледный шар) – запуск или включение ориентировочной реакции, движений конечностей, пищевого поведения (жевание, глотание и т. д.).
Дисфункции бледного шара приводят к гипомимии, гиподинамии, эмоциональной тупости, вызывают маскообразность лица, тремор головы, конечностей, миоклонию - быстрые подергивания отдельных мышечных групп, создают монотонную речь. Начало движений затрудняется, исчезают вспомогательные и реактивные движения при вставании, нарушаются содружественные движения рук при ходьбе.
Функции ограды. Ограда – узкая полоска серого вещества, медиально граничит с наружной капсулой, латерально – с капсулой экстрема. Ограда имеет прямые и обратные связи с островковой корой, а также лобной, затылочной, височной. Ограда связана с обонятельной луковицей и обонятельной корой с ипси- и контралатеральной стороны, а также с оградой другого полушария. Из подкорковых образований ограда связана со скорлупой, хвостатым ядром, черным веществом, миндалевидным комплексом, зрительным бугром, бледным шаром. Нейроны ограды участвуют в реакциях возбудительного характера на соматические, слуховые, зрительные раздражения (ориентировочные реакции, поворот головы, жевательные, глотательные, рвотные движения). При повреждении ограды левого полушария у человека наблюдается расстройство речи.
Таким образом, базальные ганглии головного мозга являются интегративными центрами организации моторики, эмоций, высшей нервной деятельности, причем каждая из этих функций может быть усилена или заторможена активацией отдельных образований базальных ядер.
Смирнов В.М., Яковлев В.Н. Физиология центральной нервной системы: Учеб. пособие для студ. высш. учеб. заведений. – М.: Академия, 2002. – с. 181 – 200
Структурно-функциональная организация лимбической системы
В более узком понимании в лимбическую систему включают образования древней коры (обонятельная луковица и бугорок), старой коры (гиппокамп, зубчатая и поясная извилины), подкорковые ядра (миндалина, ядра перегородки), и этот комплекс рассматривается по отношению к гипоталамусу и ретикулярной формации ствола как более высокий уровень интеграции вегетативных функций. В настоящее время преобладает понимание лимбической системы в более широком плане: кроме вышеназванных структур в нее также включают зоны новой коры лобной и височной долей, гипоталамус и РФ среднего мозга.
Важнейшие афферентные входы в лимбическую систему осуществляются от различных областей головного мозга, а также через гипоталамус от РФ ствола, которая считается главным источником ее возбуждения. Важным стимулирующим афферентным входом являются также импульсы от обонятельных рецепторов по волокнам обонятельного нерва, поступающие в структуры лимбической системы: обонятельные луковицу и бугорок, параобонятельную зону Брока, препириформную кору, миндалину. Главные эфферентные выходы из лимбической системы осуществляются через гипоталамус (особенно его мамиллярные тела) на нижележащие вегетативные и соматические центры ствола и спинного мозга. Другой эфферентный выход проводит возбуждение из лимбической системы в новую кору (преимущественно ассоциативную). Через нее лимбическая система включается в регуляцию высших психических функций.
Характерным свойством лимбической системы является наличие хорошо выраженных кольцевых нейронных связей, объединяющих различные ее структуры. Эти связи дают возможность длительной циркуляции (реверберации) возбуждения, которая является механизмом его пролонгирования, повышения проводимости синапсов и формирования памяти. Реверберация возбуждения создает условия для сохранения единою функционального состояния структур замкнутого крута и навязывания этого состояния другим структурам мозга. Важнейшим циклическим образованием лимбической системы является лимбический круг Пейпеца, идущий от гиппокампа через свод к мамиллярным телам, затем к передним ядрам таламуса, далее в поясную извилину и через парагиппокампову извилину обратно к гиппокампу. Этот круг играет большую роль в формировании эмоций, обучении и памяти. Другой лимбический круг (от миндалины через терминальную полоску к мамиллярным телам гипоталамуса, затем к лимбической области среднего мозга и обратно к миндалинам) имеет важное значение в формировании агрессивно-оборонительных, пищевых и сексуальных реакций.
Функции лимбической системы
Получая информацию о внешней и внутренней среде организма, лимбическая система после сравнения и обработки этой информации запускает через эфферентные выходы вегетативные, соматические и поведенческие реакции, обеспечивающие приспособление организма к внешней среде и сохранение внутренней среды на определенном уровне. В этом состоит наиболее общая функция лимбической системы.
Механизмы этого приспособления связаны с регуляцией лимбической системой висцеральных функций, в связи с чем лимбическуто систему иногда называют «висцеральным мозгом». Эта функция осуществляется преимущественно через деятельность гипоталамуса, который является диэнцефалическим звеном лимбической системы.
О тесных эфферентных связях лимбической системы (через гипоталамус) с внутренними органами свидетельствуют разнообразные изменения их функций при раздражении лимбических структур, особенно миндалин. При этом эффекты имеют различный знак в виде активации или угнетения висцеральных функций: происходит повышение или понижение частоты сердечных сокращений, моторики и секреции желудка и кишечника, секреции различных гормонов аденогипофизом (особенно АКТГ и гонадотропинов).
Важнейшей функцией лимбической системы является формирование эмоций, т. е. переживаний, в которых отражается субъективное отношение человека к предметам внешнего мира и результатам собственной деятельности. В свою очередь, эмоции являются субъективным компонентом мотиваций — состояний, запускающих и реализующих поведение, направленное на удовлетворение возникших потребностей. Через механизм эмоций лимбическая система улучшает приспособление организма к изменяющимся условиям среды.
В иерархии мозговых структур гипоталамус является критической зоной для возникновения эмоций: перерезка ствола ниже гипоталамуса выключает эмоции. В структуре эмоций выделяют собственно эмоциональные переживания и его периферические (вегетативные и соматические) проявления. Эти компоненты эмоций могут иметь относительную самостоятельность: выраженные субъективные переживания могут сопровождаться небольшими периферическим проявлениями и, напротив, при псевдоэмоциях яркие периферические проявления, например мимические и вегетативные реакции при плаче или смехе, могут протекать без существенных субъективных переживаний. Гипоталамус является структурой, ответственной преимущественно за вегетативные проявления эмоций.
Кроме гипоталамуса к структурам лимбической системы, наиболее тесно связанным с эмоциями, принадлежат поясная извилина и миндалина. Электрическая стимуляция миндалины у человека вызывает преимущественно отрицательные эмоции — страх, гнев, ярость. Напротив, двустороннее удаление миндалин в эксперименте на обезьянах резко снижает их агрессивность, повышает тревожность, неуверенность в себе. У таких животных нарушается способность оценивать информацию (особенно зрительную и слуховую), поступающую из окружающей среды, и связывать эту информацию со своим эмоциональным состоянием. В результате нарушается нормальное взаимодействие организма с окружающей средой, в том числе зоосоциальные отношения с другими особями в группе. Наряду с этим миндалина участвует в процессе сравнения конкурирующих эмоций, выделения доминирующей эмоции (и мотивации) и, следовательно, влияет на выбор поведения. Поясная извилина, имеющая многочисленные связи как с новой корой, так и со стволовыми центрами, выполняет, по мнению некоторых авторов, роль главного интегратора различных систем мозга, формирующих эмоции.
Важную роль в регуляции эмоций играет вентральная лобная кора, имеющая хорошо выраженные связи с миндалиной. Поражение лобной коры вызывает резкие нарушения эмоций у человека; характеризующиеся возникновением эмоциональной тупости, особенно изменением эмоций, возникающих при социальных отношениях людей, творчестве, и растормаживанием эмоций, связанные с удовлетворением биологических потребностей.
Следующей важной функцией лимбической системы является ее участие в формировании памяти и осуществлении обучения. Эта функция преимущественно связана с основным лимбическим кругом Пейпеца. Вместе с тем в одной из форм обучения (однократном обучении) большое значение имеет миндалина благодаря ее свойству индуцировать сильные отрицательные эмоции, способствуя быстрому и прочному формированию временной связи.
Среди структур лимбической системы, ответственных за память и обучение, выдающуюся роль играют гиппокамп и связанные с ним задние зоны лобной коры. Их деятельность совершенно необходима для консолидации памяти — перехода кратковременной памяти в долговременную. Повреждение гиппокампа у человека вызывает резкое нарушение усвоения новой информации, образования промежуточной и долговременной памяти. […]
Базальные ганглии
Базальные ганглии — это совокупность трех парных образований, расположенных в конечном мозге в основании больших полушарий: филогенетически более древней части — бледного шара (pallidum), более позднего образования — полосатого тела (stiratum) и наиболее молодой части — ограды (claustrum). Бледный шар состоит из наружного и внутреннего сегментов; полосатое тело — из хвостатого ядра (п. caudatus) и скорлупы (putamen). Ограда расположена между скорлупой и островковой корой. В функциональном отношении в базальные ганглии включают некоторые структуры промежуточного и среднего мозга: субталамические ядра и черное вещество (substantia nigra).
Функциональные связи базальных ганглиев
Возбуждающая афферентная импульсация поступает преимущественно в полосатое тело в основном из трех источников:
1) от всех областей коры прямо и через таламус;
2) от неспецифических интраламинарных ядер таламуса;
3) от черного вещества.
Среди эфферентных связей базальных ганглиев можно отметить три главных выхода:
1) от полосатого тела тормозящие пути идут к бледному шару непосредственно и с участием субталамического ядра; от бледного шара начинается самый важный эфферентный путь базальных ганглиев, идущий преимущественно в таламус, в его двигательные вентральные ядра, от них возбуждающий путь идет в двигательную кору (поля 4 и 6);
2) часть эфферентных волокон из бледного шара и полосатого тела идет к центрам ствола мозга (ретикулярная формация, красное ядро и далее в спинной мозг), а также через нижнюю оливу в мозжечок;
3) от полосатого тела тормозящие пути идут к черному веществу и после переключения — к ядрам таламуса. Оценивая связи базальных ганглиев в целом, можно отметить, что эта структура является промежуточным звеном (станцией переключения), связывающей ассоциативную и, частично, сенсорную кору с двигательной корой.
В структуре связей базальных ганглиев в последнее время выделяют несколько параллельно действующих функциональных петель, соединяющих базальные ганглии и кору больших полушарий. Скелетно-моторная петля соединяет премоторную, двигательную и соматосенсорную области коры со скорлупой базальных ганглиев, импульсация из которой идет в бледный шар и черное вещество и далее через двигательное вентролатеральное ядро возвращается в премоторную область коры (поле 6). Считают, что эта петля служит для регуляции таких параметров движения, как амплитуда, сила, направление.
Глазодвигательная петля соединяет области коры, контролирующие направление взгляда (поле 8 лобной коры и поле 7 теменной коры), с хвостатым ядром базальных ганглиев. Оттуда импульсация поступает в бледный шар и черное вещество, из которых она проецируется соответственно в ассоциативное медиодорсальное и переднее релейное вентральное ядра таламуса, а из них возвращается в лобное глазодвигательное поле 8. Эта петля участвует в регуляции, например, скачкообразных движений глаз (саккад).
Предполагается существование также сложных петель, по которым импульсация из лобных ассоциативных зон коры поступает в структуры базальных ганглиев (хвостатое ядро, бледный шар, черное вещество) и через медиодорсальное и вентральное переднее ядра таламуса возвращается в ассоциативную лобную кору. Считают, что эти петли участвуют в осуществлении высших психофизиологических функций мозга: контроле мотиваций, прогнозировании, познавательной (когнитивной) деятельности.
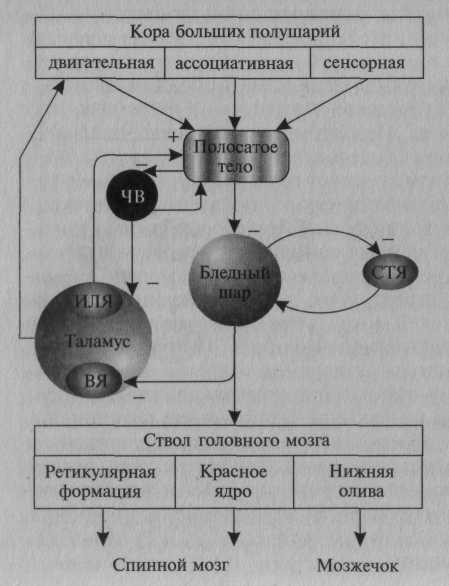
Рисунок: схема основныхафферентных и эфферентных связей базальных ганглиев:
ЧВ — черное вещество; СТЯ — субталамическое ядро; ВЯ — вентральные ядра; ИЛЯ — интраламинарное ядро; «+» — возбуждающие и «-» — тормозные влияния
Функции отдельных образований базальных ганглиев
Функции полосатого тела. Основными объектами функционального влияния полосатого тела являются бледный шар, черное вещество, таламус и моторная кора.
Влияние полосатого тела на бледный шар осуществляется преимущественно через тонкие тормозные волокна (медиатор ГАМК). Однако часть нейронов бледного шара дают смешанные ответы (ВПСП и ТПСП), а некоторые нейроны отвечают только ВПСП. Эти данные свидетельствуют о том, что полосатое тело оказывает на бледный шар двоякое действие: тормозящее и возбуждающее с преобладанием тормозящего.
Влияние полосатого тела на черное вещество. Между черным веществом и полосатым телом имеются двусторонние связи. Нейроны полосатого тела оказывают тормозящее влияние (медиатор ГАМК) на нейроны черного вещества. В свою очередь, нейроны черного вещества оказывают модулирующее влияние (медиатор дофамин) на фоновую активность нейронов полосатого тела. До сих пор окончательно не установлен характер этого влияния — тормозной, возбуждающий или тот и другой. Дофаминергическим путям в базальных ганглиях придается важное значение в связи с двигательными расстройствами типа паркинсонизма, при которых резко падает концентрация дофамина в обоих ядрах полосатого тела — хвостатом и скорлупе. Кроме влияния на полосатое тело, черное вещество оказывает тормозящее действие на нейроны таламуса (медиатор ГАМК) и получает возбуждающие афферентные входы от субталамического ядра.
Влияние полосатого тела на таламус. В середине XX в. было установлено, что раздражение участков таламуса вызывает синхронизацию электроэнцефалограммы: появление в ней высокоамплитудных ритмов, характерных для фазы медленного сна. Далее было показано, что эти ритмы можно вызвать раздражением не только таламуса, но и полосатого тела. При этом из всех структур (черное вещество, миндалина, бледный шар), посылающих прямые пути к таламусу, только раздражение полосатого тела вызывает появление в коре высокоамплитудных медленных ритмов. Напротив, разрушение полосатого тела нарушает эту реакцию и уменьшает время сна в цикле сон — бодрствование.
Влияние полосатого тела на моторную кору. В эксперименте показано тормозное влияние (медиатор ацетилхолин) хвостатого ядра полосатого тела на двигательную кору. Однако это влияние имеет не диффузный характер, а «вытормаживает» ненужные в данных условиях степени свободы движения, обеспечивая, например, четкую двигательно-оборонительную реакцию.
Прямая стимуляция полосатого тела через хронически вживленные электроды вызывает относительно простые двигательные реакции: поворот головы и туловища в сторону, противоположную раздражению, иногда сгибание конечности на противоположной стороне. Стимуляция некоторых зон полосатого тела вызывает задержку поведенческих реакций, например ориентировочной, пищедобывательной (животное словно «застывает» в одной позе). При этом на электроэнцефалограмме развиваются медленные высокоамплитудные ритмы. Раздражение других точек полосатого тела приводит к подавлению ощущения боли.
Поражение полосатого тела (его хвостатого ядра) дает характерную клиническую картину, в которой главными являются насильственные избыточные движения (гиперкинезы). Больной как бы не может справиться со своей мускулатурой. Эти гиперкинезы обозначаются как хорея и атетоз. При повреждении в эксперименте полосатого тела у млекопитающих стабильно развивается синдром гиперактивности: число бесцельных движений в пространстве увеличивается в 5 —7 раз. Опыты с разрушением полосатого тела показали его важную роль в организации поведения: животные с удаленной корой, но с сохраненными хвостатыми ядрами в упрощенном виде могут осуществлять некоторые формы поведения (передвижение в пространстве, поиск пищи и ранее выработанные простые условные рефлексы). Дополнительное разрушение хвостатых ядер прекращает эти формы поведения.
Функции бледного шара. Получая из полосатого тела преимущественно тормозное и частично возбуждающее влияния, бледный шар оказывает модулирующее влияние на двигательную кору, мозжечок, ретикулярную формацию, красное ядро. При стимуляции бледного шара у животных преобладают элементарные двигательные реакции в виде сокращения мышц конечностей, шеи и лица. При этом выявлено влияние бледного шара на некоторые зоны гипоталамуса (центр голода и задний гипоталамус), отмечена активация пищевого поведения. Разрушение бледного шара сопровождается снижением двигательной активности: возникает адинамия, как бы отвращение ко всякому движению, сонливость, эмоциональная тупость, затрудняются осуществление имеющихся и выработка новых условных рефлексов.
Подводя итог результатам деятельности базальных ганглиев, можно отметить, что участие в регуляции движения является главной, но не единственной их функцией. Наиболее важной двигательной функцией является выработка (наряду с мозжечком) сложных двигательных программ, которые реализуются через моторную кору и обеспечивают двигательный компонент поведения организма. Наряду с этим базальные ганглии контролируют такие параметры движения, как сила, амплитуда, скорость и направление. Кроме участия в регуляции движения и организации различных форм поведения базальные ганглии включаются в регуляцию цикла сон — бодрствование, в механизмы формирования условных рефлексов, в сложные формы восприятия, например осмысление текста.
Новая кора большого мозга
Новая кора (неокортекс) представляет собой слой серого вещества общей площадью 1500 — 2200 см2, покрывающий большие полушария. Она составляет около 72% всей площади коры и около 40% массы головного мозга. В коре имеется около 14 млр. нейронов, количество глиальных клеток примерно в 10 раз больше. Кора головного мозга является в филогенетическом плане наиболее молодой нервной структурой, у человека она осуществляет высшую регуляцию функций организма и психофизиологические процессы, обеспечивающие различные формы поведения.
Структурно-функциональная характеристика коры
В направлении с поверхности в глубь коры различают шесть горизонтальных слоев.
I. Молекулярный слой имеет очень мало клеток, но большое количество ветвящихся дендритов пирамидных клеток, формирующих сплетение, расположенное параллельно поверхности. На этих дендритах образуют синапсы афферентные волокна, приходящие от ассоциативных и неспецифических ядер таламуса.
II. Наружный зернистый слой составлен в основном звездчатыми клетками и частично малыми пирамидными клетками. Волокна клеток этого слоя расположены преимущественно вдоль поверхности коры, образуя кортикокортикальные связи.
III. Наружный пирамидный слой состоит преимущественно из пирамидных клеток средней величины. Аксоны этих клеток, как и зернистые клетки II слоя, образуют кортикокортикальные ассоциативные связи.
IV. Внутренний зернистый слой по характеру клеток (звездчатые клетки) и расположению их волокон аналогичен наружному зернистому слою. В этом слое афферентные волокна имеют синаптические окончания, идущие от нейронов специфических ядер таламуса и, следовательно, от рецепторов сенсорных систем.
V. Внутренний пирамидный слой образован средними и крупными пирамидными клетками, причем гигантские пирамидные клетки Беца расположены в двигательной коре. Аксоны этих клеток образуют эфферентные кортикоспинальные и кортикобульбарный двигательные пути.
VI. Слой полиморфных клеток образован преимущественно веретенообразными клетками, аксоны которых образуют кортикоталамические пути.
Оценивая в целом афферентные и эфферентные связи коры, можно отметить, что в слоях I и IV происходят восприятие и обработка поступающих в кору сигналов. Нейроны II и III слоев осуществляют кортикокортикальные ассоциативные связи. Покидающие кору эфферентные пути формируются преимущественно в V —VI слоях. Более детально деление коры на различные поля проведено на основе цитоархитектонических признаков (формы и расположения нейронов) К.Бродманом, который выделил 52 поля, многие из которых характеризуются функциональными и нейрохимическими особенностями.
Гистологические данные показывают, что элементарные нейронные цепи, участвующие в обработке информации, расположены перпендикулярно поверхности коры. Электрофизиологические исследования с погружением микроэлектродов перпендикулярно поверхности соматосенсорной коры показали, что все встречаемые на пути нейроны отвечают на раздражитель только одного качества, например тактильный. Напротив, при погружении электрода под углом на его пути попадались нейроны разной модальности. В результате был сделан вывод о наличии в коре мозга функциональных объединений нейронов, расположенных в цилиндрике диаметром 0,5 — 1,0 мм, включающем все слои коры и содержащем несколько сотен нейронов. Такие структуры были названы нейронными колонками, обнаруженными в моторной коре и различных зонах сенсорной коры. Соседние нейронные колонки могут частично перекрываться, а также взаимодействовать друг с другом по механизму латерального торможения и осуществлять саморегуляцию по типу возвратного торможения.
Кортикализация функций — возрастание в филогенезе роли коры большого мозга в анализе и регуляции функций организма и подчинение себе нижележащих отделов ЦНС. Например, регуляция таких двигательных функций, таких как локомоция (прыжки, ходьба, бег) и выпрямительные рефлексы, у низших позвоночных (амфибий) полностью обеспечивается стволом мозга, и удаление больших полушарий практически их не изменяет. У кошек перерезка ствола между средним и промежуточным мозгом существенно не влияет на выпрямительные рефлексы, но нарушает в остром периоде локомоцию, которая в дальнейшем частично восстанавливается. Выключение коры больших полушарий у обезьян и у человека приводит к потере не только локомоции, но и выпрямительных рефлексов.
Локализация функций в коре головного мозга интенсивно изучается в клинической и экспериментальной медицине, начиная с середины XIX в. При разработке этой проблемы были сформулированы две противоположные по смыслу концепции: 1) узкого локализационизма, со стремлением поместить функцию в одну отдельно взятую структуру; 2) функциональной равноценности различных корковых структур. Современная концепция локализации функций базируется на принципе многофункциональности (но не равноценности) корковьгх полей. Свойство мультифункциональности позволяет данной корковой структуре включаться в обеспечение различных форм деятельности, реализуя при этом основную, генетически присущую ей функцию. Степень мультифункциональности различных корковых структур неодинакова: например, в полях ассоциативной коры она выше, чем в первичных сенсорных полях, а в корковых структурах выше, чем в стволовых. В основе мультифункциональности лежат многоканальность поступления в кору мозга афферентного возбуждения, перекрытие афферентных возбуждений, особенно на таламическом и корковом уровнях, модулирующее влияние различных структур (неспецифического таламуса, базальных ганглиев) на корковые функции, взаимодействие корково-под корковых и межкорковых путей проведения возбуждения. Результаты изучения локализации функций позволили ученым в середине XX в. разделить кору головного мозга на различные функциональные зоны — картирование коры. Детальные функциональные карты используются в неврологической практике, однако надо иметь в виду упрощенный характер этих схем. Одним из наиболее крупных вариантов функционального разделения коры головного мозга является выделение в ней сенсорной, ассоциативной и двигательной областей.
Сенсорные области коры
Сенсорные области коры — зоны, в которые проецируются сенсорные раздражители (или: проекционная кора, корковые отделы анализаторов). Они расположены преимущественно в теменной, височной и затылочной долях. Афферентные пути в сенсорную кору поступают преимущественно от специфических сенсорных ядер таламуса (вентральных, задних латерального и медиального). Сенсорная кора имеет хорошо выраженные II и IV слои и называется гранулярной.
Зоны сенсорной коры, раздражение или разрушение которых вызывает четкие и постоянные изменения чувствительности организма, называются первичными сенсорными областями. Они состоят преимущественно из мономодальных нейронов и формируют ощущения одного качества. В первичных сенсорных зонах обычно имеется четкое пространственное (топографическое) представительство частей тела, их рецепторных полей. Вокруг первичных сенсорных зон находятся менее локализованные вторичные сенсорные зоны, полимодальные нейроны которых отвечают на действие нескольких раздражителей.
Важнейшей сенсорной областью является теменная кора постцентральной извилины и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий (поля 1 – 3), которую обозначают как соматосенсорная область. Здесь имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной чувствительности и чувствительности опорно-двигательного аппарата от мышечных, суставных, сухожильных рецепторов. Проекция участков тела в этой области характеризуется тем, что проекция головы и верхних отделов туловища расположена в нижнелатеральных участках постцентральной извилины, проекция нижней половины туловища и ног — в верхнемедиальных зонах извилины, проекция нижней части голени и стоп — в коре парацентральной дольки на медиальной поверхности полушарий. При этом проекция наиболее чувствительных участков (язык, губы, гортань, пальцы рук) имеет относительно большие зоны по сравнению с другими частями тела. Предполагается, что в зоне тактильной чувствительности языка расположена проекция и вкусовой чувствительности (первичная сенсорная зона S I).
Кроме S I выделяют меньших размеров вторичную соматосен-сорную область S II, расположенную на верхней стенке боковой борозды, на границе ее пересечения с центральной бороздой. Функции филогенетически более древней S II изучены плохо. Известно, что локализация поверхности тела в ней менее четкая, импульсация сюда поступает как от противоположной, так и от «своей» стороны, предполагают ее участие в сенсорной и моторной координации двух сторон тела.
Другой первичной сенсорной зоной является слуховая кора (поля 41, 42), которая расположена в глубине латеральной борозды (кора поперечных височных извилин Гешля). В этой зоне в ответ на раздражение слуховых рецепторов кортиева органа формируются звуковые ощущения, изменяющиеся по громкости, тону и другим качествам. Здесь имеется четкая топическая проекция: в разных участках коры представлены различные участки кортиева органа. К проекционной коре височной доли относится предположительно также центр вестибулярного анализатора в верхней и средней височных извилинах (поля 20 и 21). Обработанная сенсорная информация используется для формирования «схемы тела» и регуляции функций мозжечка (височно-мосто-мозжечковый путь).
Еще одна первичная проекционная область новой коры расположена в затылочной коре — первичная зрительная область (кора части клиновидной извилины и язычковой дольки, поле 17). Здесь имеется топическое представительство рецепторов сетчатки, и каждой точке сетчатки соответствует свой участок зрительной ' коры, при этом зона желтого пятна имеет сравнительно большую зону представительства. В связи с неполным перекрестом зрительных путей в зрительную область каждого полушария проецируются одноименные половины сетчатки. Наличие в каждом полушарии проекции сетчатки обоих глаз является основой бинокулярного зрения. Раздражение коры 17-го поля приводит к возникновению световых ощущений. Около поля 17 расположена кора вторичной зрительной области (поля 18 и 19). Нейроны этих зон полимодальны и отвечают не только на световые, но и на тактильные, слуховые раздражители. В данной зрительной области происходит, синтез различных видов чувствительности и возникают более сложные зрительные образы и их опознание. Раздражение этих полей вызывает зрительные галлюцинации, ауру (навязчивые ощущения), движение глаз.
Основная часть информации об окружающей среде и внутренней среде организма, поступившая в сенсорную кору, передается для дальнейшей ее обработки в ассоциативную кору.
Ассоциативные области коры
Ассоциативные области коры (синоним: межсенсорная, межанализаторная кора) включают участки новой коры большого мозга, которые расположены рядом с сенсорными и двигательными зонами, но не выполняют непосредственно чувствительных или двигательных функций. Границы этих областей обозначены недостаточно четко, неопределенность преимущественно связана со вторичными (высшими) проекционными зонами, функциональные свойства которых являются переходными между свойствами первичных проекционных и ассоциативных зон. Ассоциативная кора является филогенетически наиболее молодой частью новой коры, получившей наибольшее развитие у приматов и человека. У последнего она составляет около 50 % всей коры или 70 % неокортекса.
Основной физиологической особенностью нейронов ассоциативной коры, отличающей их от нейронов первичных зон, является полисенсорность (полимодальность): они отвечают с почти одинаковым порогом не на один, а на несколько раздражителей — зрительные, слуховые, кожные и др. Полисенсорность нейронов ассоциативной коры создается как ее кортикокортикальными связями с разными проекционными зонами, так и главным ее афферентным входом от ассоциативных ядер таламуса, в которых уже произошла сложная обработка информации от различных чувствительных путей. В результате этого ассоциативная кора представляет собой мощный аппарат конвергенции различных сенсорных возбуждений, позволяющий произвести сложную обработку информации о внешней и внутренней среде организма и использовать ее для осуществления высших психофизиологических функций.
По таламокортикальным проекциям выделяют две ассоциативные системы мозга: таламотеменную и таламолобную. Некоторые авторы выделяют таламовисочную систему.
Таламотеменная система представлена ассоциативными зонами теменной коры (поля 5, 7, 40), получающими основные афферентные входы от задней группы ассоциативных ядер таламуса (латеральное заднее ядро и подушка). Теменная ассоциативная кора имеет эфферентные выходы на ядра таламуса и гипоталамуса, моторную кору и ядра экстрапирамидной системы. Основными функциями таламотеменной системы являются гнозис, формирование «схемы тела» и праксис. Под гнозисом понимают функцию различных видов узнавания; формы, величины, значения предметов, понимание речи, познание процессов, закономерностей и др. К гностическим функциям относится оценка пространственных отношений, например взаимного расположения предметов. В теменной коре выделяют центр стереогнозиса, расположенный сзади от средних отделов постцентральной извилины (поля 7, 40, частично 39) и обеспечивающий способность узнавания предметов на ощупь. Вариантом гностической функции является формирование в сознании трехмерной модели тела («схемы тела»), центр которой расположен в поле 7 теменной коры. Под праксисом понимают целенаправленное действие, центр его находится в над-краевой извилине (поля 39 и 40 доминантного полушария) и обеспечивает хранение и реализацию программы двигательных автоматизированных актов (например, рукопожатие, причесывание, зажигание спички и др.).
Таламолобная система представлена ассоциативными зонами лобной коры (поля 9 – 14), имеющими основной афферентный вход от ассоциативного медиодорсального ядра таламуса. Главной функцией лобной ассоциативной коры является формирование программ целенаправленного поведения, особенно в новой для человека обстановке. Реализация этой общей функции основывается на других функциях таламолобной системы:
1) формирование доминирующей мотивации, обеспечивающей направление поведения человека. Эта функция основана на тесных двусторонних связях лобной коры с лимбической системой и ролью последней в регуляции высших эмоций человека, связанных с его социальной деятельностью и творчеством;
2) обеспечение вероятностного прогнозирования, что выражается изменением поведения в ответ на изменения обстановки окружающей среды и доминирующей мотивации;
3) самоконтроль действий путем постоянного сравнения результата действия с исходными намерениями, что связано с созданием аппарата предвидения (акцептор результата действия в функциональной системе согласно теории П.К. Анохина).
В результате проведения по медицинским показаниям префронтальной лоботомии, при которой пересекаются связи между лобной долей и таламусом, наблюдается развитие «эмоциональной тупости», отсутствие мотивации, твердых намерений и планов, основанных на прогнозировании. Такие люди становятся грубыми, нетактичными, ненадежными, у них появляется тенденция к повторению каких-либо двигательных актов, хотя обстановка уже изменилась и надо выполнять другие действия.
Концепция таламовисочной системы не получила еще достаточной научной проработки. Говоря о роли височной коры, надо отметить, что некоторые ассоциативные центры, например стереогнозиса и праксиса, включают в себя и участки височной коры (поле 39). В височной коре расположен слуховой центр речи Вернике, находящийся в задних отделах верхней височной извилины (поля 22, 37 и 42 левого доминантного полушария). Этот центр обеспечивает речевой гнозис — распознание и хранение устной речи, как собственной, так и чужой. В средней части верхней височной извилины (поле 22) находится центр распознания музыкальных звуков и их сочетаний. На границе височной, теменной и затылочной долей (поле 39) находится центр чтения письменной речи, обеспечивающий распознание и хранение образов письменной речи.
Психофизиологические функции, осуществляемые ассоциативной корой, инициируют поведение организма, обязательным компонентом которого являются произвольные целенаправленные движения, осуществляемые при обязательном участии двигательной коры.
Двигательные области коры
[…] В современной физиологии в двигательной коре выделяют первичную и вторичную моторные области.
В первичной моторной коре (прецентральная извилина, поле 4) расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела. При этом проекции мышц нижних конечностей и туловища расположены в верхних участках прецентральной извилины и занимают сравнительно небольшую площадь, а проекции мышц верхних конечностей, лица и языка расположены в нижних участках извилины и занимают большую площадь («двигательный человечек» Пенфильда). Основной закономерностью топографического представительства является то, что регуляция деятельности мышц, обеспечивающих наиболее точные и разнообразные движения (речь, письмо, мимика), требует участия больших по площади участков двигательной коры. Двигательные реакции на раздражение первичной моторной коры осуществляются с минимальным порогом (высокая возбудимость), они представлены элементарными сокращениями мышц противоположной стороны тела (для мышц головы сокращение может быть билатеральное). При поражении этой корковой зоны утрачивается способность к тонким координированным движениям конечностей, особенно пальцев рук.
Вторичная двигательная кора (поле 6) расположена на латеральной поверхности полушарий, впереди прецентральной извилины (премоторная кора). Она осуществляет высшие двигательные функции, связанные с планированием и координацией произвольных движений. Кора поля 6 получает основную часть эфферентной импульсации базальных ганглиев и мозжечка и участвует в перекодировании информации о плане сложных движений. Раздражение коры поля 6 вызывает сложные координированные движения, например поворот головы, глаз и туловища в противоположную сторону, содружественные сокращения мышц-сгибателей или мышц-разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры, связанные с социальными функциями человека: центр письменной речи в заднем отделе средней лобной извилины (поле 6), центр моторной речи Брока в заднем отделе нижней лобной извилины (поле 44), обеспечивающие речевой праксис, а также музыкальный моторный центр (поле 45), определяющий тональность речи, способность петь.
В структурном плане моторную кору называют агранулярной корой, так как в ней плохо выражены зернистые слои (II и IV); V слой, напротив, выражен лучше, чем в других зонах коры, и содержит гигантские пирамидные клетки Беца. Нейроны двигательной коры получают афферентные входы через таламус от мышечных, суставных и кожных рецепторов, а также от базальных ганглиев и мозжечка. Основной эфферентный выход двигательной коры на стволовые и спинальные моторные центры формируют пирамидные клетки V слоя. Пирамидные и сопряженные с ними вставочные нейроны расположены вертикально по отношению к поверхности коры. Такие рядом лежащие нейронные комплексы, выполняющие сходные функции, называют функциональными двигательными колонками. Пирамидные нейроны двигательной колонки могут возбуждать или тормозить мотонейроны стволовых и спинальных центров, например, иннервирующих одну мышцу. Соседние колонки в функциональном плане перекрываются, а пирамидные нейроны, регулирующие деятельность одной мышцы, расположены обычно не в одной, а в нескольких колонках.
Основные эфферентные связи двигательной коры осуществляются через пирамидные и экстрапирамидные пути, начинающиеся от гигантских пирамидных клеток Беца и менее крупных пирамидных клеток V слоя коры прецентральной извилины (60 % волокон), премоторной коры (20% волокон) и постцентральной извилины (20% волокон). Крупные пирамидные клетки имеют быстро про водящие аксоны и фоновую импульсную активность около 5 Гц, которая при движении увеличивается до 20 — 30 Гц. Эти «быстрые» пирамидные клетки иннервируют крупные (высокопороговые) α -мотонейроны в двигательных центрах ствола и спинного мозга. От мелких пирамидных клеток отходят тонкие медленнопроводяшие миелиновые аксоны. Эти клетки имеют фоновую активность около 15 Гц, которая во время движения увеличивается или уменьшается. Они иннервируют мелкие (низкопороговые) α -мотонейроны в стволовых и спинальных двигательных центрах.
Пирамидный путь состоит из 1 млн волокон кортикоспинального пути, начинающихся от коры верхней и средней трети прецентральной извилины, и 20 млн волокон кортикобульбарного пути, начинающегося от коры нижней трети прецентральной извилины (проекция лица и головы). Волокна пирамидного пути оканчиваются на α-мотонейронах двигательных ядер III – VII и IX – XII черепных нервов (кортикобульбарный путь) или на спинальных двигательных центрах (кортикоспинальный путь). Через двигательную кору и пирамидные пути осуществляются произвольные простые движения и сложные целенаправленные двигательные программы, например профессиональные навыки, формирование которых начинается в базальных ганглиях и мозжечке и заканчивается во вторичной моторной коре. Большинство волокон пирамидных путей осуществляют перекрест, однако небольшая часть волокон остается неперекрещенными, что способствует компенсации нарушенных функций движения при односторонних поражениях. Через пирамидные пути осуществляет свои функции и премоторная кора: двигательные навыки письма, поворот головы, глаз и туловища в противоположную сторону, а также речь (речедвигательный центр Брока, поле 44). В регуляции письма и особенно устной речи имеется выраженная асимметрия больших полушарий мозга: у 95% правшей и 70% левшей устная речь контролируется левым полушарием.
К корковым экстрапирамидным путям относятся кортикорубральные и кортикоретикулярные пути, начинающиеся приблизительно от тех зон, которые дают начало пирамидным путям. Волокна кортикорубралъного пути оканчиваются на нейронах красных ядер среднего мозга, от которых далее идут руброспинальные пути. Волокна кортикоретикулярных путей оканчиваются на нейронах медиальных ядер ретикулярной формации моста (от них идут медиальные ретикулоспинальные пути) и на нейронах ретикулярных гигантоклеточных ядер продолговатого мозга, от которых начинаются латеральные ретикулоспинальные пути. Через эти пути осуществляется регуляция тонуса и позы, обеспечивающих точные целенаправленные движения. Корковые экстрапирамидные пути являются компонентом экстрапирамидной системы головного мозга, к которой относятся мозжечок, базальные ганглии, моторные центры ствола; система осуществляет регуляцию тонуса, позы, координацию и коррекцию движений. Поскольку кортикопирамидные пути отдают многочисленные коллатерали структурам экстрапирамидной системы, обе системы работают в функциональном единстве, поэтому некоторые авторы считают нецелесообразным их разделение.
Оценивая в общем роль различных структур головного и спинного мозга в регуляции сложных направленных движений, можно отметить, что побуждение (мотивация) к движению создается в лимбической системе, замысел движения — в ассоциативной коре больших полушарий, программы движений — в базальных ганглиях, мозжечке и премоторной коре, а выполнение сложных движений происходит через двигательную кору, моторные центры ствола и спинного мозга.