

6.2. Репликация днк.
6.2.1. Репликация – процесс самоудвоения ДНК, или биосинтез дочерней молекулы ДНК, полностью идентичной исходной молекуле (матрице). Локализация процесса – клеточное ядро.
Условия, необходимые для репликации ДНК:

Рисунок 6.1. Репликация ДНК (объяснения в тексте).
1) Матрица – молекула ДНК (рисунок 6.1, а);
2) Расплетающие белки – разрывают водородные связи между комплементарными основаниями двойной спирали ДНК, в результате чего образуется репликативная вилка (рисунок 6.1, б);
3) Субстраты и источники энергии – дезоксирибонуклеозидтрифосфаты (дАТФ, дГТФ, дТТФ, дЦТФ). Присоединяются к азотистым основаниям полинуклеотидных цепей при помощи водородных связей по принципу комплементарности;
4) ДНК-полимераза – фермент, который формирует из нуклеозидтрифосфатов новые полинуклеотидные цепи за счёт образования 3’,5’-фосфодиэфирных связей. Синтез идёт в направлении от 5’-конца к 3’-концу. На одной ветви репликативной вилки синтезируется непрерывная цепь, на другой – фрагменты Оказаки (рисунок 6.1, в);
5) ДНК-лигаза – фермент, соединяющий фрагменты Оказаки в единую цепь (рисунок 6.1, г).
В результате образуются две идентичные молекулы ДНК (рисунок 6.1, д).
6.3. Транскрипция
6.3.1. Транскрипция – биосинтез РНК на матрице ДНК. Процесс транскрипции также происходит в клеточном ядре. Условия, необходимые для транскрипции:
Матрица – участок одной из цепей ДНК (рисунок 6.2, а);
ДНК-зависимая РНК-полимераза – главный фермент, участвующий в транскрипции. Место присоединения фермента к ДНК – промотор;
Субстраты и источники энергии – рибонуклеозидтрифосфаты (АТФ, ГТФ, УТФ, ЦТФ). Связываются с азотистыми основаниями транскрибируемой цепи ДНК водородными связями по принципу комплементарности. При участии ДНК-зависимой РНК-полимеразы образуются 3’,5’-фосфодиэфирные связи. Направление полимеризации цепи РНК – от 5’-конца к 3’-концу. Сигналом к окончанию транскрипции служат терминирующие последовательности нуклеотидов (рисунок 6.2,б).
В результате образуются первичные транскрипты – предшественники мРНК, тРНК и рРНК. Затем происходит их посттранскрипционное созревание (процессинг). Процессинг мРНК включает:
вырезание неинформативных участков – интронов (рисунок 6.2, в);
сращивание (сплайсинг) информативных участков – экзонов (рисунок 6.2, г);
защиту концевых участков от действия нуклеаз (рисунок 6.2, д):
присоединение 7-метилгуанилата («колпачок») к 5’-концу 5’,5’- фосфодиэфирной связью;
присоединение полиаденилового нуклеотида к 3’-концу.
Образующаяся мРНК поступает из ядра в цитоплазму клеток.
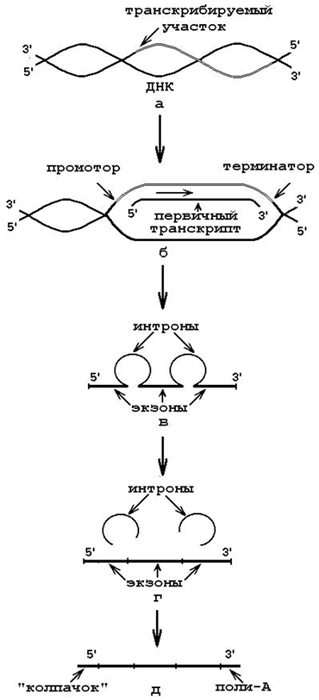
Рисунок 6.2. Транскрипция и созревание первичного транскрипта (объяснения в тексте).
6.4. Трансляция
6.4.1. Трансляция (от англ. translation – перевод) – перевод генетической информации, заключённой в мРНК, в линейную последовательность аминокислот в полипептидной цепи. Этот перевод осуществляется посредством генетического (биологического) кода.
6.4.2. Генетический код – последовательность нуклеотидов, соответствующая определённым аминокислотам. Генетический код характеризуется свойствами:
код триплетный – каждой аминокислоте соответствует тройка (триплет) нуклеотидов – кодон. Всего существует 43 = 64 кодона. Из них 61 является смысловым (то есть кодирует определённую аминокислоту) и 3 – бессмысленными (терминирующими);
код неперекрывающийся – один и тот же нуклеотид ДНК или РНК не может принадлежать одновременно двум соседним кодонам;
код непрерывный – отсутствуют «знаки препинания», вставки между кодонами в полинуклеотидной цепи;
код вырожденный (множественный) – некоторые аминокислоты могут кодироваться более, чем одним триплетом нуклеотидов (так как кодонов 61, а аминокислот – 20);
код универсальный – смысл кодонов одинаков для организмов всех видов.
6.4.3. Аминокислоты и триплеты нуклеотидов, кодирующие их, не комплементарны друг другу. Поэтому должны существовать молекулы-адапторы, каждая из которых может взаимодействовать как с определённым кодоном, так и с соответствующей аминокислотой. Такими молекулами являются транспортные РНК (рисунок 6.3). Каждая тРНК содержит триплет нуклеотидов – антикодон, который комплементарен строго определённому кодону мРНК.
3’-конец тРНК (акцепторный участок) является местом присоединения аминокислоты, соответствующей кодону мРНК.

Рисунок 6.3. Схема строения транспортной РНК.
6.4.4. Активация аминокислот – подготовительный этап биосинтеза белка – включает связывание их со специфическими тРНК при участии фермента аминоацил-тРНК-синтетазы. Реакция происходит в цитоплазме клеток. Схема этой реакции показана на рисунке 6.4.

Рисунок 6.4. Реакция активации аминокислот.
6.4.5. Собственно процесс трансляции включает 3 стадии – инициации, элонгации, терминации и происходит на рибосомах.
Каждая рибосома состоит из большой и малой субчастиц (40S и 60S) и содержит аминоацильный (А) и пептидильный (П) участки. Пептидильный участок связывает инициирующую аминоацил-тРНК, все остальные аминоацил-тРНК присоединяются к аминоацильному участку.
Стадии трансляции представлены на рисунке.

Рисунок 6.5. Стадии трансляции (объяснения в тексте).
1) Стадия инициации – начало трансляции. Условия, необходимые для инициации:
инициирующий кодон мРНК (АУГ);
белковые факторы инициации;
малая и большая субчастицы рибосомы;
ГТФ (источник энергии для смыкания субчастиц рибосомы);
ионы магния;
инициирующая аминоацил-тРНК (метионил-тРНК) – связывается своим антикодоном с инициирующим кодоном мРНК в пептидильном участке рибосомы.
В результате образуется инициирующий комплекс: мРНК – рибосома – метионил-тРНК (рисунок 6.5, а).
2) Стадия элонгации – удлинение полипептидной цепи на 1 аминокислотный остаток – происходит в три шага:
присоединение к инициирующему комплексу аминоацил-тРНК, соответствующей кодону, находящемуся в аминоацильном участке рибосомы (рисунок 6.5 ,б);
транспептидация – образование пептидной связи между остатками аминокислот (рисунок 6.5, в). Источник энергии – ГТФ;
транслокация – перемещение рибосомы относительно мРНК на 1 триплет (рисунок 6.5, г). Источник энергии – ГТФ. В ходе элонгации принимают участие белковые факторы.
Описанный процесс многократно повторяется (по количеству аминокислот в цепи).
3) Стадия терминации – окончание трансляции. Обеспечивается присутствием в цепи мРНК одного из терминирующих (бессмысленных) кодонов – УАА, УГА или УАГ. В освобождении полипептида участвуют белковые факторы терминации (рисунок 6.5, д). Когда в аминоацильном участке оказывается один из бессмысленных кодонов, факторы терминации стимулируют гидролазную активность пептидилтрансферазы. Благодаря этому гидролизуется связь между тРНК и пептидом. ГТФ для этой реакции не требуется. После этого пептидная цепь, тРНК и мРНК покидают рибосому, а её субчастицы диссоциируют.
Таким образом, трансляция мРНК приводит к формированию пептидной цепи со строго определённой последовательностью аминокислотных остатков. Следующий этап формирования белка – фолдинг, т.е. сворачивание пептидной цепи в правильную трёхмерную структуру. Если белок состоит из нескольких субъединиц, то фолдинг включает и объединение их в единую макромолекулу.
Считается, что небольшие белковые молекулы, содержащие около 100 аминоацильных остатков, могут самостоятельно принимать трёхмерную структуру, фолдинг более крупных полипептидных цепей требует участия специальных белков – шаперонов.
Посттрансляционные модификации белковой молекулы могут включать: частичный протеолиз; объединение нескольких полипептидных цепей в олигомерный белок; присоединение простетической группы; модификации аминокислотных остатков (гидроксилирование пролина в гидроксипролин в коллагене, метилирование аргинина в гистоне, йодирование тирозина в тироглобулине).
Ряд химических веществ, способных тормозить синтез белка, используется в лечении инфекционных заболеваний и злокачественных опухолей. К ним относятся многие антибиотики (см. таблицу 7).
Таблица 7