

МОРФО-ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ИММУННОЙ СИСТЕМЫ
Общая морфо-функциональная характеристика иммунной системы
Иммунная система осуществляет защиту организма от генетически чужеродных тел и веществ, называемых антигенами. Такая функция иммунной системы обеспечивается ее способностью отличать "свое" от "чужого" и обеспечивать ингибирование и элиминацию генетически чужеродных структур. Причем под генетически чужеродными структурами понимают не только посторонние организмы (вирусы, бактерии) и синтезированные ими биополимеры, но и собственные генетически измененные в результате мутаций при воспроизведении клетки организма. Следовательно, основной физиологической функцией иммунной системы является специфическая иммунологическая реактивность, реализующаяся благодаря тому, что эта система
-
концентрирует антиген
-
обеспечивает контакт с антигеном различных видов клеток
-
транспортирует гуморальные вещества и клеточные структуры лимфоидной ткани в необходимые участки организма
-
в конечном итоге элиминирует чужеродный антиген.
Иммунологическая реактивность может проявляться в виде двух типов процессов:
-
выработке антител, которые либо непосредственно уничтожают антиген, либо облегчают его уничтожение макрофагами, нейтрофилами или системой комплимента (гуморальные иммунные реакции)
-
в непосредственном повреждающем действии лимфоцитов по отношению к антигенам (клеточные иммунные реакции).
Для иммунной системы характерны следующие морфо-функциональные особенности:
-
она генерализована по всему телу
-
ее клетки постоянно рециркулируют по всему организму через лимфо- и кровоток
-
она обладает уникальной способностью вырабатывать сугубо специфические молекулы антител, различные по своей специфике в отношении каждого антигена.
В организме человека содержится примерно 1011 делящихся клеток. Если допустить, что мутации возникают в 10-5 – 10-7 случаев делений (подобно бактериям), то в организме человека при однократном воспроизведении пула пролиферирующих клеток должно образовываться примерно 105-106 мутантных клеток. В норме такие клетки или возникшие из них конгломераты ингибируются иммунной системой и элиминируются. Таким образом, иммунная система обеспечивает поддержание генетического гомеостаза в организме, а, следовательно, гомеостаза физико-химической структуры макромолекул и клеток. Дефекты специфической иммунологической реактивности приводят либо к потере способности различать "чужое" (вследствие чего может возникнуть рак и длительные инфекционные заболевания), либо утрате способности различать "свое" (что служит причиной возникновения аутоиммунных заболеваний вследствие реакции иммунной системы против собственных тканей).
Наряду со специфической иммунологической реактивностью иммунной системы, проявляющейся в ее способности распознавать "чужое" и обеспечивать нетерпимость к нему, она обладает не менее важным свойством иммунологической толерантности, заключающейся в способности обеспечивать терпимость к "своему".
Органы иммунной системы расположены в различных частях организма и анатомически обособлены друг от друга; их основа образована лимфоидной или миелоидной тканью. Миелоидная ткань (от греч. миелос – костный мозг) располагается в мозгу, находящемся в полостях костей (костном мозгу), и является местом образования эритроцитов, кровяных пластинок, гранулоцитов, моноцитов и В-лимфоцитов (т.е. почти всех форменных элементов крови за исключением Т-лимфоцитов). Лимфоидная ткань получила свое название в связи с очень большим изобилием лимфоцитов, в ней находящихся; она образует тимус, селезенку, лимфатические узлы и неинкапсулированные лимфоидные скопления в рыхлой волокнистой соединительной ткани слизистых оболочек и кожи.
В кроветворных органах (красном костном мозге, образованном миелоидной тканью, и тимусе, образованном лимфоидной тканью) в ячеях ретикулярной ткани расположены созревающие клетки крови и их предшественники.
В основе лимфоидных органов лежит ретикулярная паренхима, образованная в большинстве случаев ретикулярной тканью (а в тимусе – ретикулярной и эпителиальной тканью) и имеющая вид сети, в ячеях которой находятся лимфоциты, являющиеся главными функциональными элементами лимфоидной ткани. Лимфоидные органы составляют 5% от массы тела человека (общая их масса составляет в среднем 1,5-2 кг), а общее число лимфоидных клеток – 1012. Лимфоциты (Т- и В-лимфоциты) представляют собой иммунологически компетентные клетки, служащие ключевым звеном:
-
в реакциях гиперчувствительности замедленного типа и трансплантационного иммунитета (формы клеточного иммунитета),
-
предшественниками антителообразующих клеток (плазматических клеток), обеспечивающих осуществление гуморальных иммунных реакций
-
носителями иммунологической памяти
-
кроме того, лимфоциты выделяют ряд гуморальных факторов, вовлекающих в иммунный ответ иммунокомпетентные и вспомогательные клетки, вступают в кооперативное взаимодействие с ними.
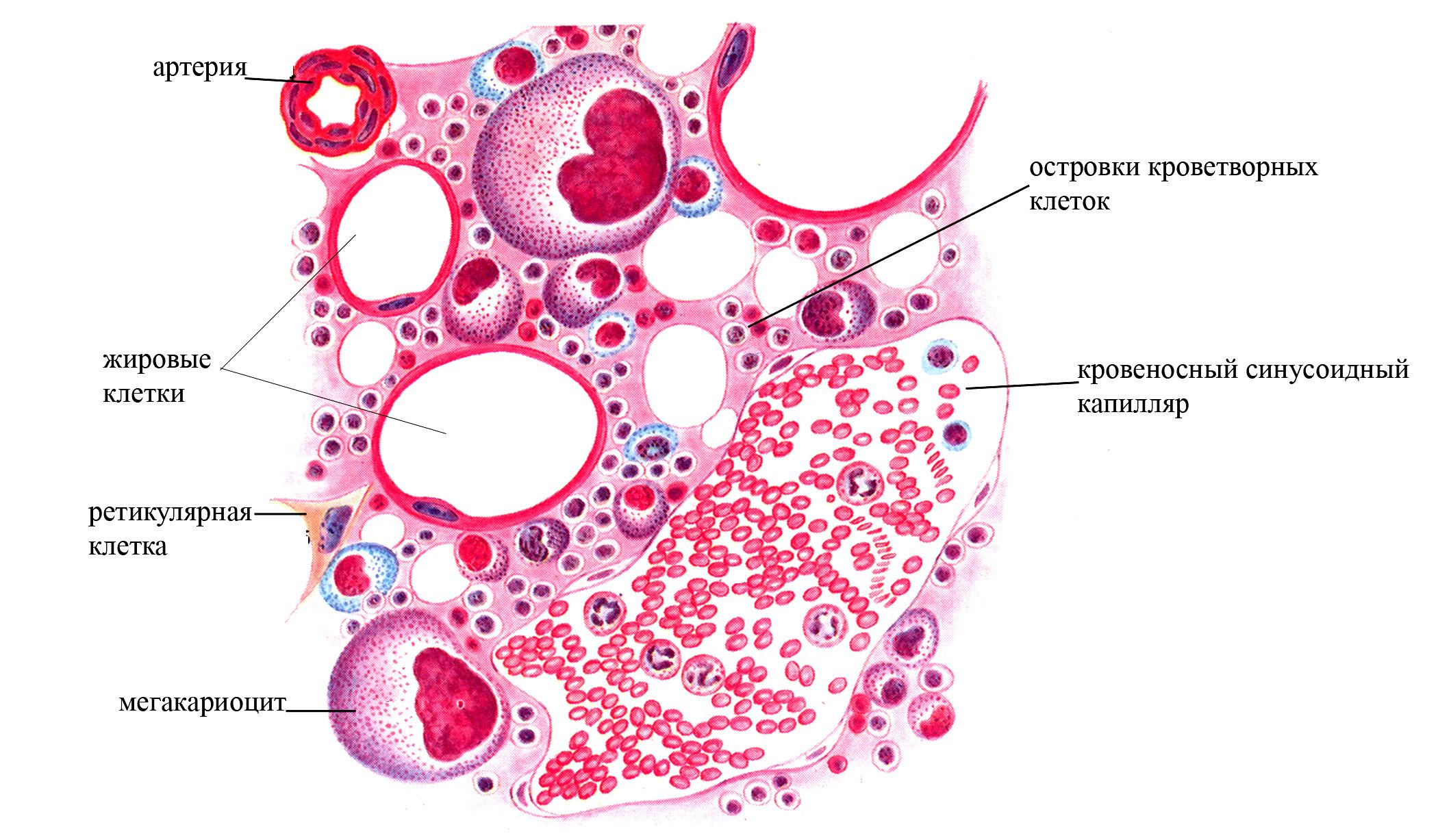
 А
А
Б
Рис. 1 Красный костный мозг. А – схема строения. Б – микрофотография красного костного мозга (окраска гематоксилин-эозином)

Рис. 2. Синусоидный капилляр красного костного мозга (вид при сканирующей электронной микроскопии)
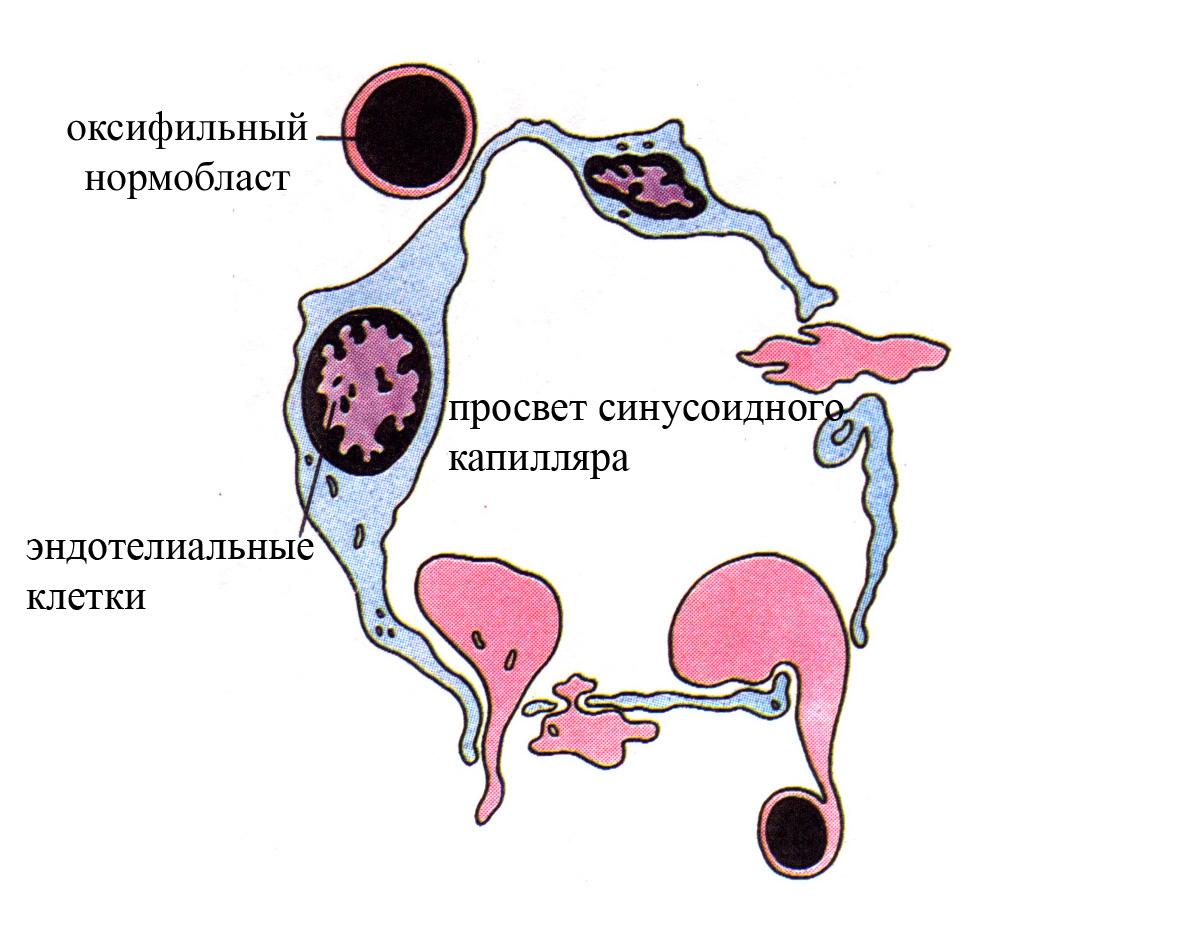
Рис. 3. Схема, демонстрирующая выход эритроцитов через стенку синусоидного капилляра красного костного мозга в кровоток

Рис. 4. Эпителиальная ткань тимуса (вид при сканирующей электронной микроскопии)
Органы иммунной системы в зависимости от их функционального назначения классифицируют на:
-
центральные органы иммунной системы – органы, в которых осуществляется лимфоцитопоэз, служат своеобразной "фабрикой" по производству лимфоцитов. Следовательно, центральные органы иммунной системы, являясь местом лимфоцитопоэза, сообщают иммунологическую компетентность клеткам-предшественникам и регулируют иммунологическую реактивность всей иммунной системы посредством вырабатываемых ими гуморальных факторов. Причем сами эти органы не осуществляют активного иммунного ответа, они заселяются стволовыми клетками и являются местом усиленного лимфопоэза даже в эмбриональном периоде и у безмикробных животных, когда периферические лимфоидные органы остаются неактивными. Удаление центральных органов иммунной системы приводит к нарушению функций иммунитета, резкому снижению числа лимфоцитов в периферической крови и периферических лимфоидных органах. Эти органы представлены:
-
красным костным мозгом (образован миелоидной тканью). У взрослого человека располагается в полостях губчатого костного вещества костей свода черепа, ребер и грудины, телах позвонков, губчатых частях некоторых коротких костей и в эпифизах длинных трубчатых костей. Наряду с эритроцитами, кровяными пластинками, гранулоцитами и моноцитами, служит местом образования В-лимфоцитов. Причем на долю лимфоцитов в красном костном мозге приходится 10% от всех ядерных его клеток. Кроветворение именно в красном костном мозге обусловлено тем, что он заложен в полостях костной ткани, которые не могут бесконечно расширяться (в отличие, например, от волокнистых соединительных тканей), и это заставляет образующиеся зрелые форменные элементы крови переходить в капилляры. Причем для костного мозга характерны расширенные капилляры с прерывистым эндотелием (синусоидные капилляры), что облегчает переход в них эритроцитов и других форменных элементов. Сами же синусоидные капилляры костного мозга не могут спадаться под действием окружающих их форменных элементов благодаря высокому гидростатическому давлению в них, обусловленному тем, что приносящие артериолы, ответвляющиеся от костных артерий, шире венул. Такая особенность микроциркуляторного русла, обеспечивающая повышенное гидростатическое давление в капиллярах, в других местах организма приводила бы к отеку, который не возникает в костной ткани благодаря большой плотности кости.
-
тимусом (вилочковая железа), лимфоидный орган, является местом образования Т-лимфоцитов. Причем все форменные элементы крови происходят от общего родоначального предшественника – стволовой клетки (называемой также колониеобразующей единицей – КОЕ). Эта стволовая клетка или, возможно, ее потомки с током крови из красного костного мозга приносятся в тимус, где дают начало Т-лимфоцитам.
-
-
периферические органы иммунной системы – органы, в которых в отличие от центральных органов иммунной системы не происходит первичного лимфоцитопоэза (т.е. антигеннезависимого лимфоцитопоэза из кроветворных стволовых клеток). Периферические органы иммунной системы заселяются лимфоцитами и служат местом контакта лимфоцитов с антигенами, т.е. представляют собой плацдарм для осуществления иммунных реакций (преимущественно гуморального типа), в результате которых происходит элиминация антигенов. К периферическим органам иммунной системы относят:
-
лимфатические узлы, где осуществляется фильтрация лимфы, оттекающей из тканей и ее очищение от различных антигенов (бактерий, вирусов, их белковых макромолекул). Лимфа образуется в тканях путем фильтрации тканевой жидкости в слепо начинающиеся в тканях лимфатические капилляры. Причем особенностью лимфатических капилляров, в отличие от кровеносных, является отсутствие базальной мембраны, что облегчает прохождение в них крупномолекулярных белков, воспалительных экссудатов и даже отдельных клеток из тканей. Лимфатические капилляры сливаются друг с другом, переходя в более крупные сосуды, выносящие лимфу из органов. Лимфатические сосуды, прежде чем доставить лимфу в кровоток, обязательно проходят через лимфатические узлы. Причем они входят в лимфатические узлы со стороны выпуклой их поверхности, а выходят – со стороны вогнутой. Лимфа, выходящая из лимфатического узла, обогащена лимфоцитами (в основном Т-лимфоцитами) и частично очищена от антигенов. Лимфатические сосуды, прошедшие через лимфатические узлы, по ходу сливаются друг с другом, образуя все более крупные сосуды. Наконец, лимфа через грудной и правый лимфатические протоки впадает в верхнюю полую вену (возвращается в кровь). Таким образом, система лимфообращения непрерывно выносит лимфу в венозную часть системы кровообращения, так что вся избыточная жидкость, формирующаяся в организме, возвращается в кровяное русло, проходя на своем пути лимфатические узлы.
-
селезенка, выступает в качестве депо крови, принимая участие в очистке ее от различных антигенов, обеспечивает элиминацию старых эритроцитов
-
неинкапсулированная лимфоидная ткань, заложенная в рыхлой соединительной ткани слизистых оболочек и кожи, обеспечивает очистку от антигенов преимущественно тканевой жидкости.
-
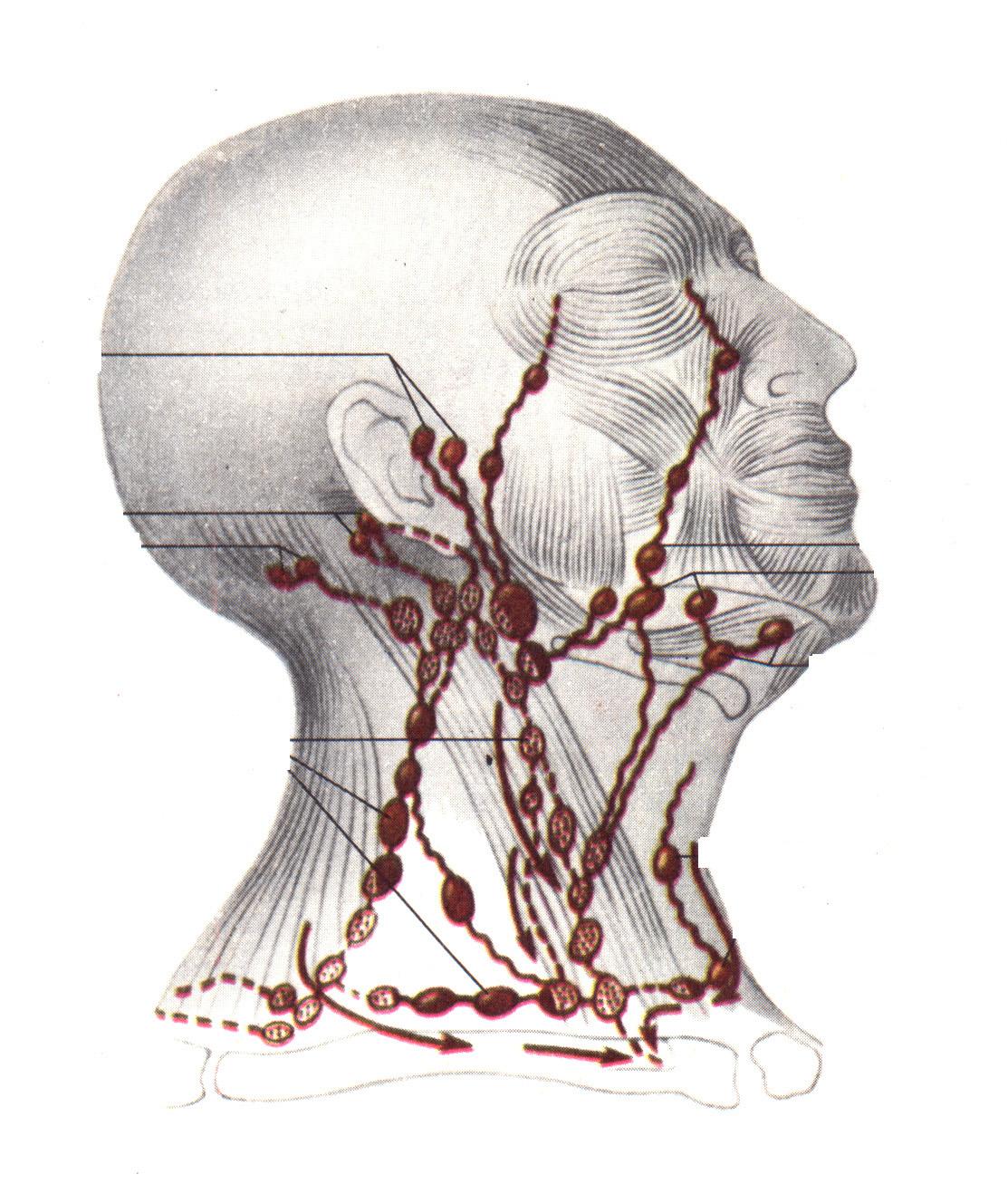
Р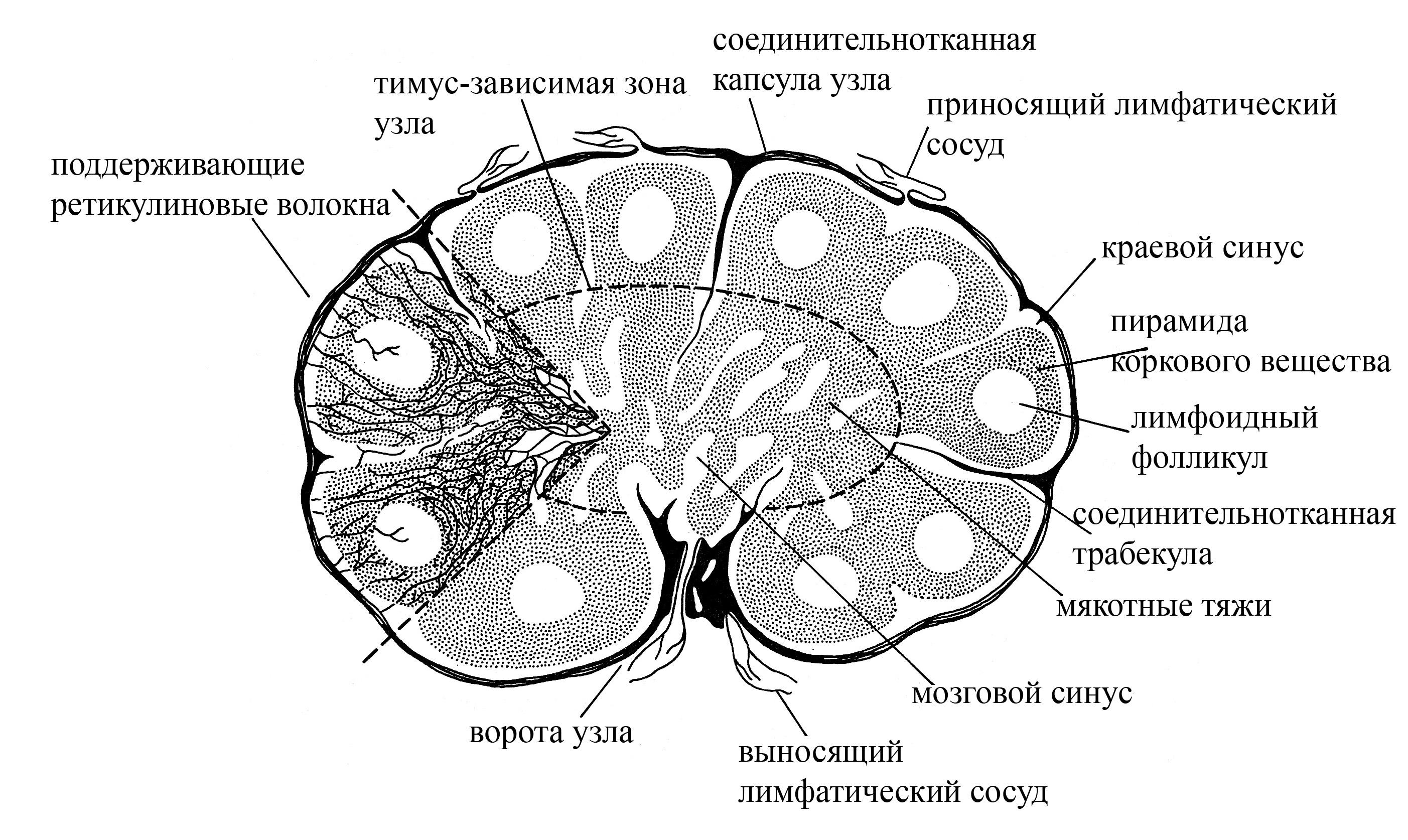 ис.
5. Лимфатические узлы и сосуды головы и
шеи человека
ис.
5. Лимфатические узлы и сосуды головы и
шеи человека
Рис. 6. Схема строения лимфатического узла. Обратите внимание на то, что приносящие лимфатические сосуды входят в лимфатический узел со стороны наружной (выпуклой) его поверхности. После чего лимфа изливается в краевой лимфатический синус, расположенный под соединительнотканной капсулой узла. Из краевого лимфатического синуса лимфа поступает в промежуточные синусы коркового вещества, откуда переходит в мозговые синусы, открывающиеся в выносящие лимфатические сосуды, которые выходят из узла в области ворот. Таким образом, лимфа фильтруется через лимфатический узел и, проходя по его синусам, постоянно контактирует с лимфоцитами и макрофагами, находящимися в лимфатических синусах, что обеспечивает очистку лимфы от различных антигенов.

Рис. 7. Неинкапсулированный лимфоидный фолликул рыхлой волокнистой соединительной ткани
Лимфатические узлы и неинкапсулированные лимфоидные скопления расположены в большинстве своем на путях проникновения инфекционных агентов в организм (в области кожи, слизистых оболочек), а, следовательно, выполняют защитно-барьерную функцию. Причем между периферическими органами иммунной системы существует "своеобразное разделение труда": они специализируется на контакте с антигенами определенных компонентов внутренней среды организма. Так, неинкапсулированные лимфоидные фолликулы контактируют преимущественно с антигенами, которые оказываются в тканевой жидкости, лимфатические узлы эффективно взаимодействуют с лимфой, протекающей через них. Наконец, в селезенке осуществляется активный и весьма длительный контакт иммуннокомпетентных клеток с антигенами, находящимися в крови, протекающей через селезенку.
Особенностью иммунной системы является ее адаптивная направленность, позволяющая обеспечить иммунный ответ на любые теоретически возможные чужеродные макромолекулы и клетки, которые могли бы проникнуть во внутреннюю среду организма. Данное свойство иммунной системы обусловлено тем, что значительное количество клеток, отвечающих специфически на данный антиген, образуется только после специфического антигенного стимула (т.е. после контакта с каким-то антигеном) и, будучи индуцированы, эти клетки выполняют свои специфические функции автономно. Следовательно, приобретение лимфоцитами иммунокомпетентности происходит не в центральных иммунных органах, где осуществляется лимфоцитопоэз, а в периферических иммунных органах после контакта с теми или иными антигенами.
Иммунная система по организации и механизмам функционирования сходна с нервной системой. Сходство заключается в том, что функции обеих этих систем генетически детерминированы, обе они представлены центральными и периферическими органами, способны реагировать на различные сигналы, имеют огромное количество рецепторных структур, специфическую память. Основным же функциональным отличием иммунной системы от нервной является то, что иммунная система способна к передаче и генерализации специфических сигналов мигрирующим клеткам (лимфоцитам), тогда как нервная – неподвижным клеткам.
Морфо-функциональная характеристика тимуса
Тимус (вилочковая железа) представляет собой лимфоидный орган, расположенный большей частью в грудной клетке, непосредственно позади рукоятки грудины, в межплевральной области, спереди перикарда. Тимус состоит из двух долей, соединенных между собой и образующих единое целое, имеет удлиненную уплощенную треугольную форму, розовато-серый цвет, вершина его направлена в область шеи.
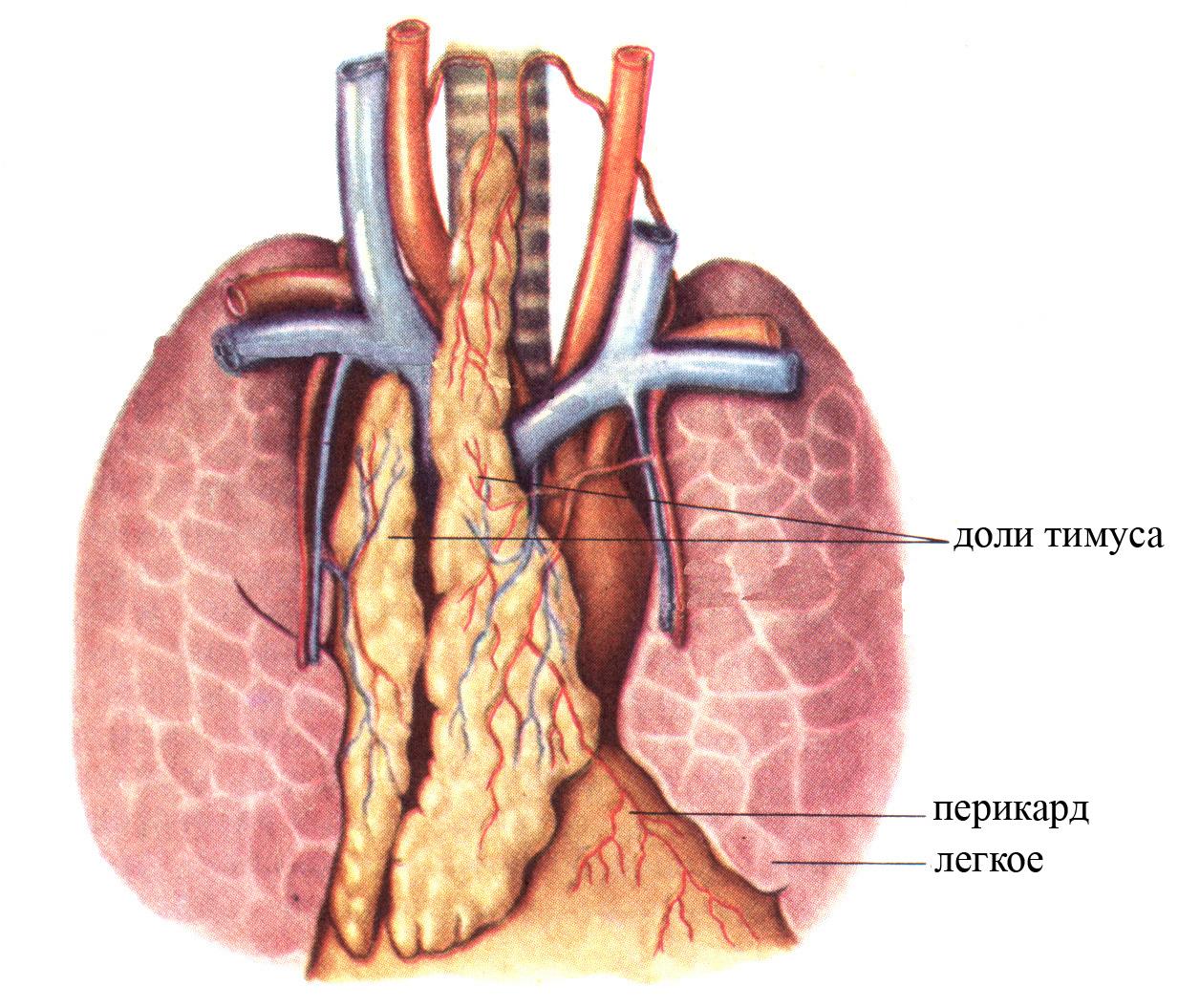
Рис. 8. Тимус (общий вид).
Вилочковая железа энтодермального происхождения. Ее эпителиальный компонент начинает закладываться в конце 1-го - начале 2-го месяца внутриутробной жизни из 3-го глоточного кармана в виде парных тяжей, которые в дальнейшем утрачивают связь с глоткой, опускаются в грудную полость и срастаются между собой. Мезенхима, окружающая эпителиальные зачатки тимуса, дает начало его соединительнотканной капсуле и многочисленным ее производным – септам, формирующим в совокупности соединительнотканную строму органа. Причем соединительнотканные септы тимуса, отходящие от его капсулы, не обеспечивают полного разделения паренхимы тимуса на дольки (орган имеет псевдодольчатое строение). Вскоре между эпителиальными клетками тимуса начинают появляться лимфоциты, которые дифференцируются либо из самой кроветворной стволовой клетки либо из ее потомков – предшественников лимфоцитов, мигрирующих из красного костного мозга. Лимфоциты становятся главными клетками тимуса. Таким образом, в тимусе можно выделить три гистологических компонента:
-
эпителиальный, являющийся производным энтодермы глоточного кармана,
-
соединительнотканный, имеющий мезенхимное происхождение и формирующий соединительнотканную капсулу и строму органа,
-
лимфоидный, представленный лимфоидными клетками на разных стадиях дифференцировки, которые являются производными стволовой кроветворной клетки, мигрирующей из красного костного мозга.
Тимус формируется раньше других органов иммунной системы и к моменту рождения имеет значительную массу (10-15 г), в течение первых 3-ех лет постнатальной жизни тимус растет наиболее интенсивно, а затем его масса и размеры относительно стабилизируются (достигая 30-40 г) и остаются таковыми до окончания периода пубертации (в среднем до 20 лет), после чего начинается возрастная инволюция тимуса (в пожилом возрасте масса органа уменьшается до 13-15 г). Причем возрастная инволюция тимуса сопровождается не просто уменьшением его массы, но и определенными морфологическими изменениями. В частности, в органе увеличивается доля соединительной и жировой ткани (у людей старше 50 лет она составляет 90% от массы органа), значительно сокращается объем эпителиальной паренхимы и лимфоидного компонента.
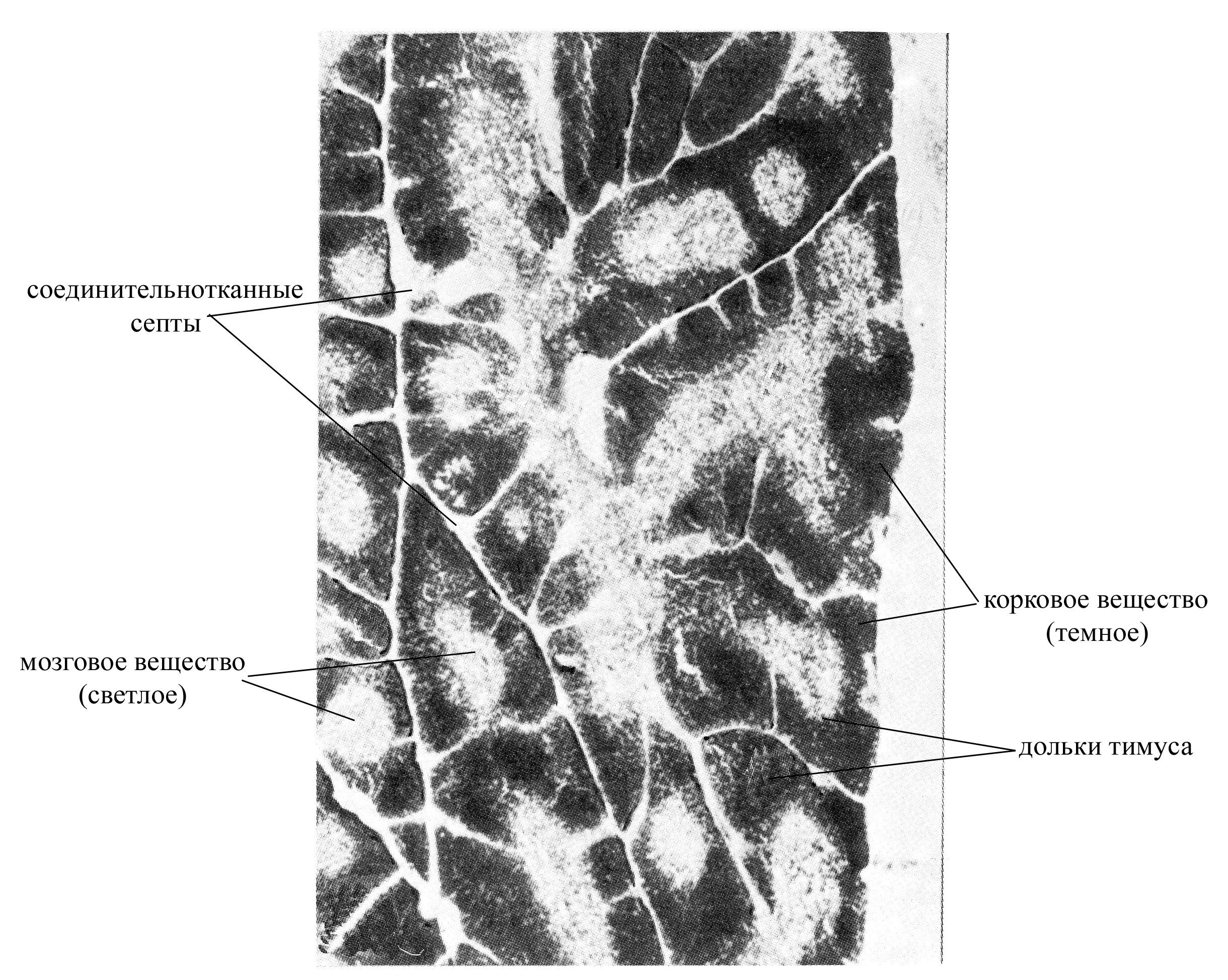
Рис. 9. Микрофотография тимуса при малом увеличении
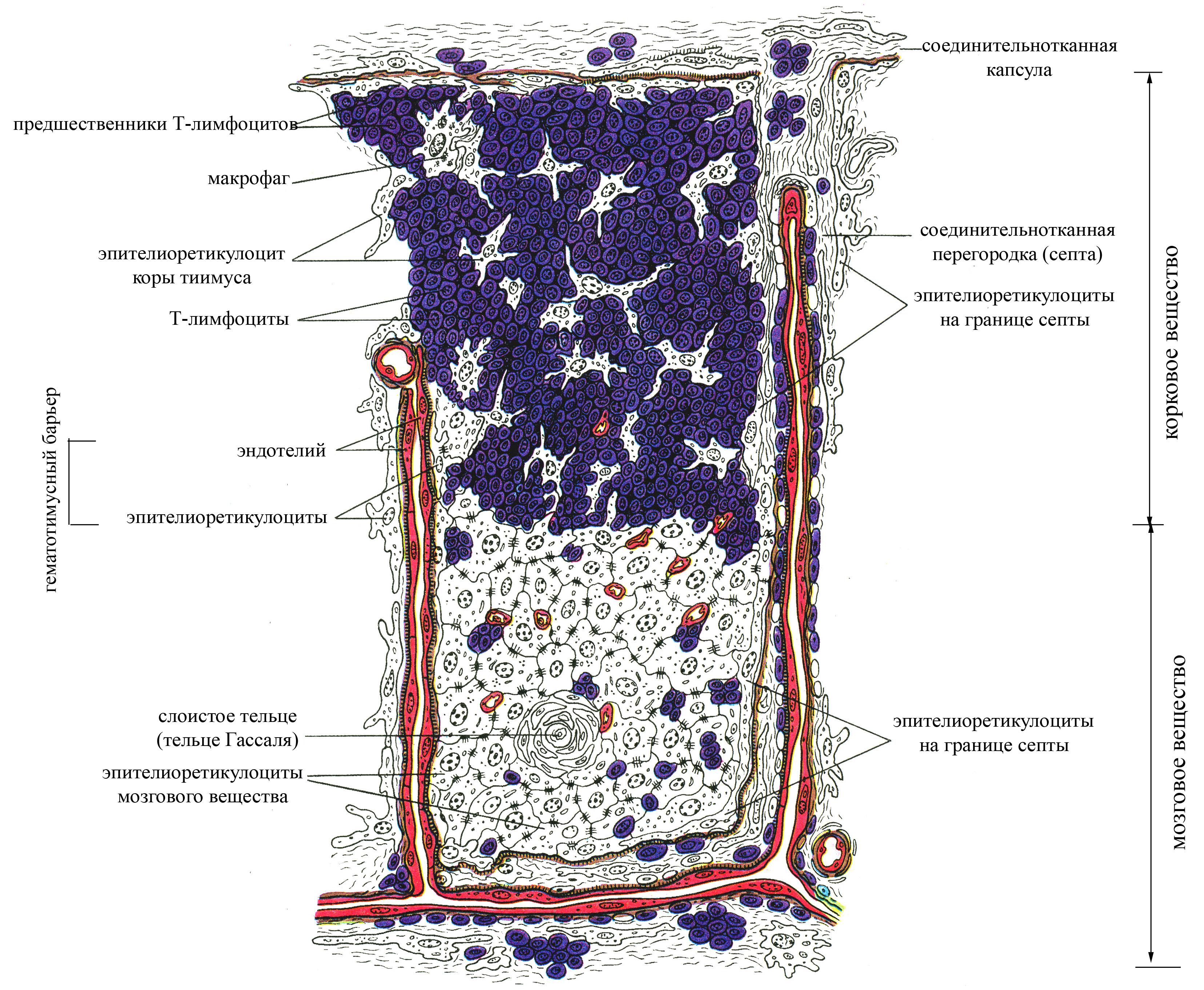
Рис. 10. Структурная организация дольки тимуса
В паренхиме тимусе различают:
-
корковое вещество, располагается непосредственно под капсулой органа, имеет более плотную консистенцию, образовано эпителиальными древовидными клетками, контактирующими отростками и формирующими таким образом сеть. В ячеях этой сети располагается большое количество лимфоцитов на разных стадиях дифференцировки. Причем самое периферическое положение в тимусе занимают наиболее крупные и наименее дифференцированные лимфоидные клетки – лимфобласты, для которых характерны частые митозы (эти клетки располагаются ближе всего к капсуле тимуса). По мере созревания размеры лимфоидных клеток уменьшаются, сокращается их митотическая активность, и они перемещаются ближе к центральной части тимуса (т.е. в направлении мозгового вещества). Таким образом, корковое вещество тимуса является автономной "фабрикой" Т-лимфоцитов, где особенно в эмбриональный и ранний постнатальный период осуществляется интенсивное их образование. Т-лимфоциты, образующиеся в корковом веществе тимуса, в дальнейшем заселяют другие лимфоидные органы, в которых превращаются в окончательно зрелые формы. Ежедневное количество новообразованных в корковом веществе Т-лимфоцитов, выходящих из тимуса, очень велико и приблизительно в 4 раза превышает нормальную общую популяцию лимфоцитов крови. Эпителиальные древовидные клетки в корковом веществе тимуса со всех сторон окружают развивающиеся лимфоидные клетки, принимая участие в создании здесь гематотимического барьера, образованного
-
эндотелиальными клетками капилляров с окружающей их базальной мембранной,
-
периваскулярной соединительной тканью, содержащей тканевую жидкость и макрофаги,
-
отростками эпителиальных древовидных клеток коркового вещества тимуса.
-

Рис. 11. Ультраструктура гематотимического барьера в корковом веществе тимуса (вид при электронной микроскопии)
Благодаря гематотимическому барьеру в корковом веществе тимуса исключается возможный контакт развивающихся лимфоцитов с антигенами внутренней среды организма, приносимыми кровью. Важным для развивающихся в корковом веществе Т-лимфоцитов является и тот факт, что в капсулу тимуса, в отличие от лимфатического узла не входят лимфатические сосуды, в результате чего лимфа не проходит через тимус, и это также исключает возможность доставки в корковое вещество органа чужеродных антигенов. Ранний контакт Т-лимфоцитов, коммитированных на какой-то антиген, с этим антигеном в корковом веществе тимуса сопровождается уничтожением такого клона Т-лимфоцитов. Благодаря уничтожению определенных клонов Т-лимфоцитов на уровне коркового вещества тимуса непосредственно после их возникновения и еще до момента их расселения в другие лимфоидные органы становится возможным выработка иммунологической толерантности по отношению к собственным антигенам гистосовместимости, широко представленным на поверхности эпителиальных древовидных клеток, окружающих развивающиеся лимфоидные клетки в корковом веществе. Вместе с тем наличие гематотимического барьера в корковом веществе тимуса, исключающее возможность контакта чужеродных антигенов, циркулирующих в крови, с развивающимися лимфоидными клетками, препятствует уничтожению клонов Т-лимфоцитов, коммитированных на чужеродные антигены и формированию иммунологической толерантности по отношению к чужеродным антигенам. В опытах на животных показано, что введение непосредственно в корковое вещество тимуса какого-то чужеродного антигена в эмбриональный период еще до момента расселения Т-лимфоцитов из тимуса в другие лимфоидные органы (селезенку и лимфатические узлы), сопровождается формированием иммунологической толерантности по отношению к этому антигену (т.е. такой антиген не воспринимается организмом как чужеродный и не вызывает иммунных реакций). Около 95% вновь образующихся в корковом веществе тимуса Т-лимфоцитов погибает, поскольку они вероятнее всего коммитированы на собственные антигены тканей организма, и лишь 5% лимфоцитов поступает в общий кровоток и далее оседает в лимфатических узлах и селезенке, где они могут контактировать с чужеродными антигенами. Часть зрелых Т-лимфоцитов из коркового вещества тимуса мигрирует в мозговое с током венозной крови, оттекающей от коркового вещества благодаря определенным особенностям кровоснабжения тимуса. В частности, артериолы тимуса проходят на границе коркового и мозгового вещества и дают начало самостоятельным капиллярным сетям этих двух зон тимуса. Вместе с тем венозная кровь, оттекающая от коркового вещества, собирается в венулы и вены мозгового, благодаря чему обязательно проходит через мозговое вещество тимуса. Кроме того, в мозговое вещество тимуса Т-лимфоциты могут приноситься с артериальной кровью, поступающей в орган (рециркулирующие полностью зрелые Т-лимфоциты). Каждый образующийся в корковом веществе тимуса Т-лимфоцит заранее коммитирован на какой-то определенный антиген (на плазматической мембране зрелых Т-лимфоцитов расположены рецепторы, которые могут избирательно взаимодействовать лишь с каким-то строго специфичным антигеном).
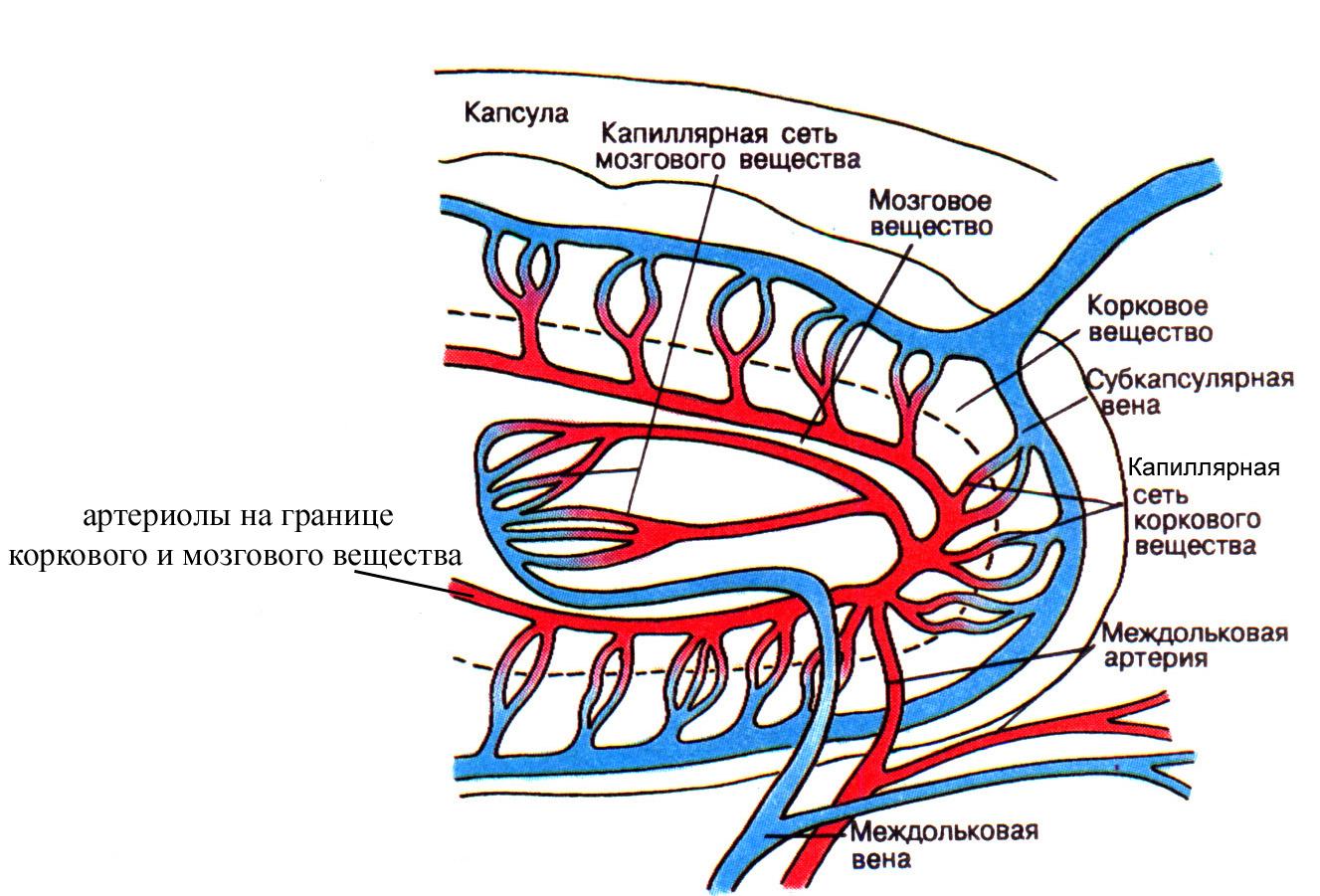
Рис. 12. Схема кровоснабжения дольки тимуса. Обратите внимание, что артериолы не проникают вглубь коркового вещества тимуса, а проходят, как правило, на границе коркового и мозгового вещества, разветвляясь на капиллярные сети коркового и мозгового вещества.
-
мозговое вещество, занимает центральное положение в тимусе, характеризуется более рыхлой консистенцией, образовано сетью древовидных эпителиальных клеток и расположенных между ними малых лимфоцитов (зрелых форм лимфоцитов). Для мозгового вещества тимуса характерна меньшая плотность лимфоцитов. Эпителиальная паренхима мозгового вещества тимуса представлена двумя компонентами:
-
диффузно расположенными древовидными эпителиоцитами, формирующими сеть, подобную таковой коркового вещества,
-
плотно упакованными эпителиоцитами, образующими слоистые тельца в мозговом веществе (называемые тельцами Гассаля), округлой формы, диаметром до 100 мкм; в таких тельцах многочисленные слои эпителиальных клеток располагаются вокруг одной центральной клетки тельца. Появляются эти тельца в тимусе на 12-ой неделе эмбрионального развития, их количество достигает максимума к 11 годам, а с 15 лет начинает уменьшаться. Роль этих телец до конца не ясна. Было замечено, что после введения в организм антигенов в тельцах происходят определенные циклические изменения и обнаруживается антиген. Существует предположение, согласно которому клетки, образующие тельца Гассаля, обладают фагоцитарной способностью.
-
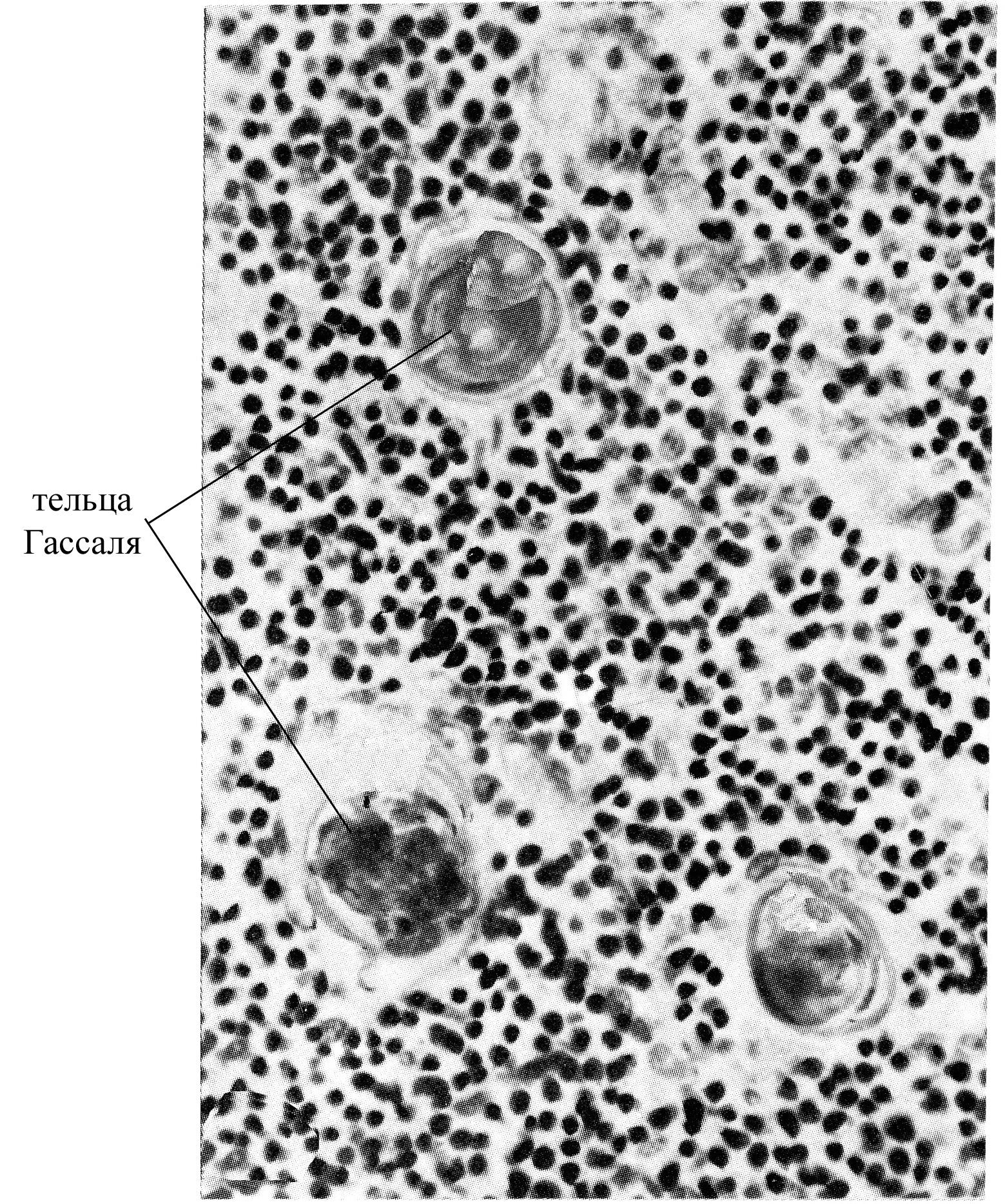
Рис. 13. Микрофотография участка мозгового вещества тимуса при большом увеличении (видны слоистые тельца или тельца Гассаля)
Эпителиальная паренхима тимуса, наряду с опорной функцией и созданием определенного микроокружения (представленного собственными антигенами гистосовместимости) вокруг Т-лимфоцитов, выполняет и эндокринную функцию. В частности, эпителиальные клетки тимуса синтезируют пептидные гормоны – тимозины и тимопоэтин. Функции тимозинов сводятся к следующему:
-
способствуют дифференцировке Т-лимфоцитов и появлению специфических рецепторов в их плазматической мембране (т.е. способствует развитию специфичности Т-лимфоцитов в отношении определенных антигенов),
-
стимулируют выработку многих цитотоксических гормонов лейкоцитами (т.н. лимфокинов, в том числе интерлейкина-2 Т-хелперами)
-
стимулируют продукцию иммуноглобулинов В-лимфоцитами и плазматическими клетками.
Другой гормон тимуса – тимопоэтин – является стимулятором дифференцировки предшественников Т-лимфоцитов, способствует дифференцировке самих Т-лимфоцитов, но в отличие от тимозинов, не влияет на развитие специфичности Т-лимфоцитов (т.е. на их коммитированность в отношении определенных антигенов).