

Физиология Karpovskyi
.pdfповторювань поєднаної дії умовного та безумовного подразників, тим міцнішим буде тимчасовий зв’язок між відповідними нервовими центрами у корі великих півкуль головного мозку. І ось настає той час, коли на вмикання лампочки (умовний подразник) без підкріплення кормом у тварини виділяється
Рис. 4. Механізм утворення умовних рефлекторних зв’язків.
А – шлях безумовного слиновидільного рефлексу; Б – зв’язок органу слуху з корою головного мозку; В – шляхи розповсюдження умовного сигналу та безумовного травного подразника; Г – шлях умовного слиновидільного рефлексу.
1 – язик; 2 – довгастий мозок; 3 – корковий безумовний травний центр; 4 – слинна залоза; 5 – звоник; 6 – вухо; 7 – слухова область кори головного мозку з тимчасовим умовно-рефлекторним центром слиновиділення; 8 – слина
слина. Утворився умовний слиновидільний рефлекс на спалахи світла. Рефлекторна дуга цього рефлексу включає: рецептори сітківки ока оптичний нерв зоровий центр кори мозку, потім новим щойно утвореним тимчасовим шляхом до центру травлення кори імпульси збудження надходять до слиновидільного центру довгастого мозку та еферентними нервовими волокнами прямують до слинних залоз, які починають виділяти слину. Таким чином, механізм утворення умовного рефлексу зводиться до утворення тимчасового зв’язку між двома корковими центрами умовного та безумовного рефлексів.
Відомий фізіолог П.К. Анохін вважає, що за умов поєднання умовного та безумовного подразників у аксоплазмі нейронів утворюються нові білкові молекули, особливо чутливі до нервових імпульсів. Ці молекули “зберігають” зв язки, що утворилися між двома корковими центрами.
351
Слід відзначити, що рефлекторні дуги умовного та безумовного рефлексів мають різний початок та загальний відцентрований шлях. Це є доказом того, що умовні рефлекси можуть утворюватися тільки на базі безумовних. Характер умовного рефлексу залежить не від умовного подразника, а від того, на базі якого безумовного рефлексу він утворений.
У даний час вважається, що в утворенні умовного рефлексу важливу роль відіграють підкоркові структури, лімбічна система та ретикулярна формація мозку. Відомо, що перелічені структури тісно поєднані з нервовими центрами кори великих півкуль головного мозку і підтримують тонус кортикальних нейронів.
КЛАСИФІКАЦІЯ УМОВНИХ РЕФЛЕКСІВ
Для утворення умовного рефлексу сигналом може слугувати будь-який індиферентний подразник. Залежно від характеру умовного подразника розрізняють натуральні та штучні умовні рефлекси.
Під час утворення натуральних умовних рефлексів умовним сигналом є будь-яка ознака, що поєднана безпосередньо з безумовним подразником, є характеристикою останнього. Наприклад, слиновиділення на вигляд, запах корму є натуральним умовним рефлексом.
Штучні умовні рефлекси утворюються за дії подразника, що не має прямого стосунку з безумовним подразником, а лише співпадає з ним за часом дії. Наприклад, у тварин можна утворити слиновидільний рефлекс на звучання метроному, дзвінка, спалахування лампи, тобто на індиферентні подразники, що підсилюється надаванням корму. Такі умовні рефлекси називають штучними.
Вже відзначалося, що умовні рефлекси утворюються на базі природжених безумовних. Такі рефлекси називають умовними рефлексами першого порядку.
І.П. Павлов зазначав, що будь-який безумовний рефлекс може слугувати базою для утворення умовних рефлексів. Немає жодної функції організму, яка не була б у змозі змінитися під впливом умовних рефлексів. Тому, регуляція більшості фізіологічних функцій здійснюється за механізмом складно-рефлекторних реакцій.
Умовні рефлекси можуть утворюватися не тільки на базі безумовних, але й на базі утвореного умовного рефлексу. Це – умовний рефлекс другого порядку. Наприклад, у собаки утворений стійкий слиновидільний умовний рефлекс на спалах лампи, коли на вмикання лампочки, без підкріплення їжею, виділяється слина (рефлекс першого порядку). Потім для утворення умовного рефлексу другого порядку використовують ще один індиферентний подразник
– звучання метронома, поєднуючи його зі спалахуванням лампочки без застосування безумовного подразника (корму). Після багаторазового поєднання звуку метронома буде спостерігатися слиновиділення без підсилення його спалахуванням лампочки. Це умовний слиновидільний рефлекс другого порядку.
З використанням рухово-захисної методики у собак можна було утворювати й умовний рефлекс третього порядку. Більш високого порядку
352
умовні рефлекси можуть утворюватися лише у людини з її розвиненою корою великих півкуль головного мозку.
ВІДМІННОСТІ УМОВНОГО РЕФЛЕКСУ ВІД БЕЗУМОВНОГО
Діяльність окремих органів, систем органів та всього організму перебуває під контролем вищого відділу центральної нервової системи – кори великих півкуль головного мозку. У механізмах регуляції цих функцій беруть участь як безумовні, так і умовні рефлекси, тобто здійснюється складно-рефлекторна реакція. Між цими двома рефлекторними актами є певні відмінності:
–безумовні рефлекси є природженими, а умовні – набутими. Під час подразнення рецепторів ротової порожнини виділяється слина – це безумовнорефлекторний акт, а виділення слини на вигляд або запах корму – умовнорефлекторна реакція;
–безумовні рефлекси видові, притаманні усім особинам даного виду. Так, усі коти вигинають спину під час появи небезпеки, собаки – гавкають, їжаки – згортаються в клубок. У тварин одного виду безумовні рефлекси є однаковими;
–умовні рефлекси індивідуальні, можуть бути неоднаковими у різних особин одного виду залежно від умов їх існування та типу вищої нервової діяльності. Кожна тварина по своєму реагує на зміну довкілля, набуваючи свій життєвий досвід;
–безумовні рефлекси виявляються за умов дії специфічних подразників на певні рецептивні поля, в той час як умовні рефлекси виникають під час дії індиферентних подразників на різноманітні рецептори різних рецептивних полів;
–безумовні рефлекси мають незмінну рефлекторну дугу та є відносно простішими, тоді як умовні рефлекси тимчасові, не мають сталої рефлекторної дуги, виникають за певних умов і зникають під час зміни цих умов;
–безумовні рефлекси можуть здійснюватися за участі нервових центрів нижчих відділів центральної нервової системи, без участі коркових центрів, тоді як у здійсненні умовних рефлексів у вищих тварин неодмінно бере участь кора великих півкуль головного мозку. При повному видаленні кори великих півкуль головного мозку втрачаються умовні рефлекси, що були утворені раніше та унеможливлюється утворення нових умовних рефлексів. Безумовнорефлекторні реакції організму при цьому зберігаються;
–безумовні рефлекси передаються у спадок, тоді як умовні рефлекси можуть закріплюватися у спадок лише за умови, що протягом декількох поколінь зовнішні подразники залишаються незмінними та утворені при цьому рефлекси корисні для тварини.
ВИДИ ГАЛЬМУВАННЯ У КОРІ МОЗКУ
Рефлекторна діяльність центральної нервової системи характеризується процесами збудження та гальмування. Збудження спричиняє здійснення умовно-рефлекторної реакції, а гальмування – пригнічує її. Умовні рефлекси утворюються за певних умов та зникають під час зміни цих умов.
Розрізняють два види гальмування умовних рефлексів: безумовне або
353
природжене та умовне або внутрішнє.
Безумовне гальмування умовних рефлексів виникає відразу при дії зовніщнього подразника, що спричиняє збудження інших нервових центрів у корі великих півкуль головного мозку. Це збудження за механізмом негативної індукції спричиняє гальмування у центрі умовного рефлексу.
Безумовне гальмування має два різновиди: зовнішнє та позамежне. Зовнішнє гальмування виникає при дії незвичайного подразника ззовні. При цьому в корі мозку виникає новий осередок збудження. Цей збуджений центр є зовнішнім стосовно центра умовного рефлексу.
Зовнішнє гальмування може бути згасаючим та таким, що не згасає. Наприклад, у собаки утворений слиновидільний рефлекс на дію умовного подразника – дзвінка. Якщо під час досліду пролунає сильний шум або з’явиться незнайома собаці людина тощо, то слиновидільний рефлекс загальмується, слина виділятися не буде. Але якщо ця людина почне з’являтися й далі під час досліду, то вона вже не буде зумовлювати гальмівну дію, щодо слиновидільного рефлексу. У цьому випадку мова йтиме про згасаюче гальмування умовного рефлексу.
Якщо під час прояву умовного рефлексу тварині нанести больове подразнення, то умовний рефлекс загальмується. І надалі це больове подразнення буде спричиняти гальмування умовного рефлексу. Це приклад зовнішнього гальмування, що не згасає.
Позамежне або охоронне гальмування умовних рефлексів виникає при підвищенні сили або тривалості дії подразника, що перевищує межі працездатності нервових клітин. Наприклад, у тварини утворений слиновидільний рефлекс на звук дзвінка. Але якщо різко збільшувати його гучність, то слиновиділення загальмується, оскільки сильний дзвінок перевищує межі працездатності нейронів. Саме таке гальмування можна спостерігати і при тривалій дії дзвінка, навіть середньої сили.
Позамежне гальмування умовних рефлексів може виникати також при дії умовних подразників не дуже великої сили, якщо нервові клітини мають низьку працездатність у результаті захворювання, втоми та інше.
Позамежне гальмування є захисним механізмом, що охороняє нервові клітини від перенапруження, тому його й називають охоронним.
Слід зазначити, що безумовне гальмування притаманне не тільки нейронам кори головного мозку, але й іншим ділянкам центральної нервової системи.
Умовне або внутрішнє гальмування притаманне лише нейронам великих півкуль головного мозку. Цей вид гальмування зароджується усередині центра умовного рефлекса, у тих самих нервових клітинах, що і збудження. Умовне гальмування виникає поступово і може тривалий час зберігатися.
Розрізняють декілька різновидів умовного гальмування: згасання, диференціювання, умовне гальмо, запізнювання.
Згасальне гальмування розвивається при систематичному непідкріпленні умовного сигналу безумовним подразником: перестає функціонувати тимчасовий зв’язок у корі мозку між нервовими центрами.
354
Якщо після повного згасання умовного рефлексу зробити перерву, то через деякий час умовний рефлекс поновлюється, деколи навіть без жодного підсилення умовного сигналу безумовним подразником.
Диференціювальне гальмування умовних рефлексів реєструється при дії іншого умовного подразника такого самого походження, на який був утворений умовний рефлекс, але відмінний за деякими характеристиками від умовного сигналу. Наприклад, у собаки утворений слиновидільний рефлекс на звук метронома з частотою 90 за хвилину. Потім вмикають метроном з частотою 120 за хвилину, який не підкріплюють кормом. Слина виділяється на звук метронома з частотою 120 за хвилину, хоча це й не підкріплюється кормом. Пояснюється це генералізацією звукового подразника, що збуджує слуховий центр у корі великих півкуль головного мозку. Надалі ж, оскільки корм підкріплюється лише звуком метронома з частотою 90 за хвилину і не підсилюється безумовним подразником 120 ударів за хвилину, слина виділяється лише на звук метронома з частотою 90 за хвилину, у якій би послідовності не вмикали метроном.
В основі такого виду гальмування лежить аналітико-синтетична діяльність нейронів кори великих півкуль головного мозку. Завдяки диференціювальному гальмуванню тварина може аналізувати різні подразники, адекватно реагувати на зміни зовнішнього середовища.
Умовне гальмо. Цей вид гальмування розвивається у тих випадках, коли до умовного сигналу, на який вже утворений умовний рефлекс приєднується інший умовний подразник, який не підкріплюється безумовним. Так, якщо у собаки утворений слиновидільний рефлекс на звук метронома, а потім до звуку метронома додають ще вмикання лампочки, то на поєднану взаємодію звуку метронома та вмикання лампочки слина не виділяється, тобто другий умовний подразник – вмикання лампочки, чинить гальмівну дію. В основі цього виду гальмування також лежить аналітико-синтетична діяльність нервових клітин кори мозку.
Запізнювання спостерігається у тих випадках, коли підкріплення безумовним подразником відбувається дещо пізніше від дії умовного сигналу (запізнюється). Якщо після дзвінка корм тварині дають через 3–4 хвилини, то умовний слиновидільний рефлекс також буде проявлятися із запізненням на такий самий час.
БІОЛОГІЧНЕ ЗНАЧЕННЯ УМОВНИХ РЕФЛЕКСІВ
Організм не може існувати без зовнішнього середовища, що підтримує його існування, оскільки із зовнішнього середовища організм одержує всі поживні речовини та оксиген, який конче необхідний для окиснення метаболітів у тканинах. Зовнішнє середовище є дуже мінливим щодо своїх параметрів. Це й викликає відповідні зміни також і в організмі тварин.
Організм тварини як саморегулююча система, вступає у взаємозв’язок із довкіллям і утворює механізми, що спрямовані на підтримання життєдіяльності в умовах мінливого зовнішнього середовища.
Ті природжені механізми реакції організму на дію факторів зовнішнього
355
середовища у формі безумовно-рефлекторних реакцій не в змозі забезпечити відповідну взаємодію організму із довкіллям та пристосування його до умов зовнішньої дійсності, що змінюється.
Слід зазначити, що умовний рефлекс ніби готує організм тварини до зустрічі з безумовним подразником. Так у тварини, що відчула запах корму, умовно-рефлекторним шляхом виділяється слина та інші травні соки і при потраплянні корму до ротової порожнини травна система вже готова до травлення. Або, заєць, почувши шурхіт у кущах, тікає, щоб не зустрітися з вовком або іншим хижаком.
Таким чином, тільки шляхом реагування на фактори довкілля та утворення умовно-рефлекторних зв’язків у корі великих півкуль головного мозку організм здатний пристосовуватися до зовнішнього середовища та підтримувати свою життєдіяльність. За допомогою умовних рефлексів тварина набуває життєвого досвіду, який дозволяє добувати корм за виглядом і запахом, рятуватися від небезпеки тощо. Тому умовні рефлекси треба розглядати, з біологічної точки зору, як форму пристосування організму до умов довкілля, що змінюються.
ДИНАМІЧНИЙ СТЕРЕОТИП
У процесі життєдіяльності організм тварини відчуває вплив комплексу подразників, які діють у певній послідовності. На кожний подразник утворюється відповідна рефлекторна реакція за участі кори великих півкуль головного мозку. Але якщо багато днів підряд діють багаточисельні подразники у незмінній послідовності та з певними проміжками часу, то у корі великих півкуль головного мозку тварини формується ціла система рефлекторних зв’язків, до того ж у тій суворій послідовності, з якою діяли подразники. Іншими словами, стереотипна дія багаточисельних подразників викликає відповідну реакцію або динамічний стереотип, що жорстко закріпилися.
Динамічний стереотип включає цілу систему цінних рефлекторних реакцій щодо багаточисельних подразників. Перший сигнал запускає усю систему рефлекторних реакцій, утворюючи цілу мозаїку осередків збудження та гальмування. Оскільки підвищується збудливість усіх нервових структур, що беруть участь у цій динамічно стереотипній реакції організму, не потрібно великих витрат енергії та напруги для здійснення багаточисельних реакцій організму. Цим диктується необхідність чіткого розкладу доби та технологічних процесів у тваринництві.
Утворення динамічного стереотипу має зв’язок із більшою напругою нервової діяльності нижчих та вищих ділянок центральної нервової системи, аналізом та синтезом найбагатшої інформації, що надходить до кори великих півкуль головного мозку. Підтримання ж установленого стереотипу не потребує великих зусиль та здійснюється з мінімальними витратами енергії.
Порушення стереотипу потребує чималих зусиль з боку нейронів кори великих півкуль головного мозку та організму в цілому, оскільки потрібно перебудувати напрацьовану систему рефлекторних взаємовідносин між нервовими центрами. Іноді перебудова стійкого динамічного стереотипу стає
356
неможливою, спостерігаються розлади вищої нервової діяльності тварини. Тому порушення динамічного стереотипу завжди негативно відбивається на здоров’ї та продуктивних якостях тварини. Це необхідно обов’язково враховувати тваринникам.
ІНСТИНКТИ
Під інстинктом розуміють сукупність безумовних рефлекторних реакцій ланцюгового характеру, коли завершення одного рефлекторного акту спонукає наступну реакцію. Інстинкти бувають різні: статеві, батьківські, харчові, побудова житла тваринами, утворення табуна та переліт птахів тощо. Збудження нервових центрів, які беруть участь у цих ланцюгових безумовнорефлекторних реакціях здійснюється як рефлекторно, починаючи з подразнення відповідних рецепторів, так і гуморально, за участі гормонів. Наприклад у механізмі статевого інстинкту беруть участь зорові рецептори, що сберуть особу протилежної статі, нюхові рецептори, чутливі до феромонів, статеві гормони, що підвищують збудливість нервових центрів, які контролюють статеві рефлекси. Або, наприклад, харчові інстинкти, що вмикають харчові рефлекси, які мають зв’язок з пошуками корму, споживання та використання поживних речовин. У мотивації пошуку та споживання корму має значення ступінь наповнення шлунка кормом, а також концентрація метаболітів у крові.
В усіх безумовно-рефлекторних реакціях, що визначають інстинкт, беруть участь нейрони різних ділянок центральної нервової системи, включаючи ретикулярну формацію, підкоркові утворення, лімбічну систему та кору великих півкуль головного мозку. Подібно будь-якому безумовнорефлекторному акту, інстинкт обростає низкою умовних рефлексів, та являє собою складно-рефлекторний акт. У тварин, позбавлених кори великих півкуль, інстинкти проявляються не в повному обсязі. Голубка без кори великих півкуль не реагує на туркотіння голуба, вона байдужа до пищання пташенят.
Інстинкти як природжена форма реакцій організму на зміну у довкіллі, забезпечує не тільки існування на певному етапі розвитку тварини, але й його ріст та подальший розвиток. Якщо ж інстинкти перестали відповідати умовам довкілля, вони можуть зникати. Але це відбувається за дуже великі проміжки часу протягом існування багатьох генерацій.
СОН ТА ГІПНОЗ
Нервові клітини кори великих півкуль головного мозку відрізняються високим рівнем обмінних процесів та відносно швидкою втомою. Відновлення працездатності нервових клітин відбувається у періоди, коли нервові клітини відмежовані від багаточисельних подразнень, коли у корі головного мозку немає потужних осередків збудження. Таким періодом є сон.
Сон – особлива діяльність мозку. Його не можна зрівнювати зі станом спокою, або гальмування. За І.П. Павловим “сон – це розлите гальмування у корі великих півкуль головного мозку зі збереженням стану бадьорості окремих ділянок, так званих “сторожових” пунктів”. Насправді, під час сну знижується функціональна активність органів дихання, кровообігу, травлення, знижується
357
збудливість тканин, тонус м’язів.
Сон є універсальним явищем. Воно спостерігається не тільки у вищих, але й у нижчих тварин. У коней та жуйних сон є періодичним та багаторазовим, тривалістю до 6 годин. Найбільш глибокий сон уночі. Більшість тварин сплять лежачи, а кінь спить переважно стоячи.
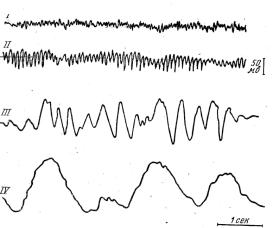
Електрофізіологічні дослідження кори великих півкуль головного мозку з реєстрацією біострумів від різих ділянок кори мозку дозволили виявити, що в організмі, який не спить, та сплячого відрізняються електроенцефалограми
(рис. 5).
|
Для тварини, що не спить, характерна |
|||||
|
наявність у електроенцефалограмі (ЕЕГ) |
|||||
|
швидких хвиль (14–40 Гц) з невеликою |
|||||
|
амплітудою (до 15 мкВ), так звані бета- |
|||||
|
ритми. У дрімотної тварини, швидкі хвилі |
|||||
|
в ЕЕГ змінюються повільними (1–3 Гц – |
|||||
|
дельта-ритми та 4–6 Гц – тета-ритми). Під |
|||||
|
час пробудження повільні хвилі різко |
|||||
|
обриваються |
та |
змінюються |
знову |
||
|
швидкими бета-ритмами. |
|
|
|||
|
Якщо подразнювати через вживлені у |
|||||
|
мозок електроди |
ретикулярну формацію |
||||
|
мозку, то характерні для сну повільні |
|||||
Рис. 5. Головні ритми ЕЕГ: |
потенціали |
згасають |
та |
змінюються |
||
І – β-ритм, ІІ – α-ритм, ІІІ – θ-ритм, |
швидкими. Таким чином, нервові імпульси, |
|||||
IV – δ-ритм |
що надходять до кори з ретикулярної |
|||||
|
формації підтримують |
високу |
активність |
|||
коркових нейронів, завдяки чому тварина стає бадьорою.
При ушкодженні ретикулярної формації у ділянці середнього мозку тварина занурюється у глибокий сон. Парентеральне застосування аміназину викликає сонливий стан у бичків, оскільки блокуються адренергічні структури ретикулярної формації і коркові нейрони не одержують активуючих сигналів.
Виділяють 5 стадій, що настають одна за одною протягом сну:
1 стадія – перехід від бадьорості до першого легкого сну. На ЕЕГ реєструються нерівномірні зиґзаґи.
2 стадія – через декілька хвилин починається нормальний сон, на ЕЕГ з’являються “сонні веретена”.
3 стадія – приблизно за 30 хвилин починається перший глибокий сон, з’являються повільні дельта-хвилі.
4стадія – за 50–70 хвилин, як тварина заснула, дельта-хвилі вщухають на короткий час (від 5 до 20 хвилин) та змінюються швидкими хвилями малої амплітуди, такими самими, що виникають під час пробудження. Але тварина не пробуджується, сон у неї глибокий. Таку стадію назвали парадоксальною.
5стадія – у подальшому парадоксальна стадія сну зникає та знову змінюється повільними хвилями (дельта-ритми).
358
Протягом 6–8-годинного сну “повільний сон” замінюється парадоксальною фазою кожні 80–90 хвилин, до того ж вона стає все більш тривалою в міру наближення пробудження.
Таким чином, можна говорити про дві фази сну:
1)сон з повільною електричною активністю, “повільний сон” з дельта-ритмами;
2)сон із швидкою електричною активністю, швидкий або “парадоксальний сон”, що виникає періодично (бета-ритми).
Виявилося, що парадоксальна фаза супроводжується рухами вух, хвоста, облизуванням, посіпуванням кінцівок, прискоренням дихання, змінами частоти серцевих скорочень, підсиленням активності мозку та підвищенням обміну речовин, але тонус більшості м’язів залишається максимально низьким.
Вчені стверджують, що всі ці зміни під час парадоксальної фази сну мають зв’язок зі сновидіннями. Сни може бачити не тільки людина, але й тварини. Вважають, що людина може розповісти свій сон лише в тому випадку, коли просинається або їй штучно переривають сон під час стадії парадоксального сну.
Отже, сон за своєю сутністю та фізіологічним механізмом не є єдиним станом. На фоні повільного сну (на ЕЕГ це дельта-хвилі) реєструються швидкі хвилі – бета-ритми (швидкий або парадоксальний сон).
У даний час вважають, що сновидіння є життєво необхідними для людей. При відсутності сновидінь можуть розвиватися різні психічні порушення, неврози та інше.
Гіпноз за І.П. Павловим це неповний сон, або сон з частковою бадьорістю. В основі гіпнозу, так само, як і сну, лежить явище іррадіації гальмування. За умов глибокого сну гальмування охоплює більшу частину кори, підкоркові утворення і навіть середній мозок, а за умов гіпнозу гальмування розповсюджується на окремі ділянки мозку. На загальному гальмівному тлі у корі великих півкуль головного мозку зберігаються осередки бадьорості, що функціонально мають зв’язок переважно зі слуховим аналізатором. Саме це дозволяє гіпнотизеру підтримувати зв’язок із тим, кого він гіпнотизує, словесним навіюванням, що має назву “рапорт”.
Гіпнотичний сон розрізняють за глибиною. У залежності від цього виділяють 3 стадії гіпнозу:
–малий – поверхневий або летаргічний;
–середній – каталептичний;
–глибокий – сомнамбулічний.
Під час поверхневого гіпнозу знижується тонус м’язів. Піддослідний чує гіпнотизера, але навіюваність ще досить мала, пробудження затруднене.
Під час середнього або каталептичного гіпнозу людина не в змозі самостійно рухатися, у свідомості з’являються “провали”. У цій стадії гіпнозу спостерігається “воскова гнучкість”.
Під час цієї ж стадії гіпнозу у деяких піддослідних (каталептиків) замість воскової гнучкості виявляється усталена напруженість усіх м’язів, тобто контрактура, каталептичний міст, коли піддослідний лежить між двох стільців,
359
спираючись на них лише потилицею та п’ятами.
У глибокому або сомнамбулічному гіпнозі свідомість піддослідного вже цілком загальмована, тіло втрачає больову та температурну чутливість.
ДВІ СИГНАЛЬНІ СИСТЕМИ ДІЙСНОСТІ
Існують дві сигнальні системи довкілля. Перша сигнальна система включає ті багаточисельні рецептори (зорові, слухові, нюхові, тактильні та інші), за допомогою яких сберуться усі зміни довкілля з наступним аналізом та синтезом інформації у нейронах кори великих півкуль головного мозку. У відповідь на всі ці подразнення організм відповідає безумовними та умовними рефлексами, що формуються та реалізуються у тварин і людей за такими самими фізіологічними механізмами.
Перша сигнальна система є загальною для тварин і людини.
Друга сигнальна система притаманна тільки людині. Це високо досконала система сигналізації, що має зв’язок із словесною сигналізацією мовою. Слова, що вимовляють, чутні та видимі, стали сигналами перших сигналів. “Якщо наші чуття та уявлення, писав І.П. Павлов, що належать до довкілля, є для нас перші сигнали дійсності, конкретні сигнали, то мова, та подразнення, що прямують до кори від мовних органів, є другі сигнали, сигнали сигналів”. Словесним сигналом людина позначає усе те, що вона сбере за допомогою свої рецепторів. Слово, як “сигнал сигналів” дає можливість відволіктися, абстрагуватися від конкретних предметів та явищ, узагальнювати інформацію, що надходить до кори мозку.
Друга сигнальна система є соціально детермінованою. Слово, мова засоби спілкування людей. Поза суспільством, без спілкування з іншими людьми, друга сигнальна система не розвивається.
Слово сбереться людиною не тільки як окремий звук або сума звуків, а як певне поняття – сбереться його змістове значення.
Жодна тварина, навіть людиноподібна мавпа, не в змозі адекватно реагувати на складні словесні подразнення. Тварина сбере інформацію з довкілля лише через першу сигнальну систему.
Особливістю другої сигнальної системи є більша швидкість утворення умовних зв язків у корі великих півкуль головного мозку.
Під час навчання утворюються багаточисельні зв’язки між різними ділянками кори мозку та центрами, що беруть участь у актах читання та письма. Так, мовні сигнали слугують підсиленням умовних подразників.
На основі другої сигнальної системи став можливим розвиток науки як вищої форми пізнання довкілля.
ТИПИ ВИЩОЇ НЕРВОВОЇ ДІЯЛЬНОСТІ
Раніше уже відзначалося, що основною формою діяльності нервової системи організму є рефлекс. У будь-якому рефлекторному акті беруть участь нейрони кори великих півкуль головного мозку, на рівні яких замикаються як безумовно-, так і умовно-рефлекторні реакції організму.
360