

Физиология Karpovskyi
.pdfлише невелика частина Т3 і Т4 залишається в крові в активному стані, а в тканинах ці комплекси розпадаються, звільнюючи тироксин і трийодтиронін. Для утворення цих гормонів необхідні амінокислота тирозин і йод.
Тиреоїдні гормони мають широкий спектр дії. Їх загальні ефекти полягають у дії на різні обмінні процеси, ріст і розвиток організму, а також в участі в адаптивних реакціях. При здійсненні пристосувальних змін в організмі тиреоїдні гормони діють спільно з іншими гормонами, перш за все глюкокортикоїдами. Найбільш виражений вплив Т3 і Т4 на енергетичний обмін. При відсутності тиреоїдних гормонів швидкість обмінних процесів знижується. При їх надлишку основний обмін у спокої може підвищуватись майже удвічі в порівнянні з нормою. Дія Т3 і Т4 полягає також в підвищенні чутливості тканин до катехоламінів; вони підсилюють глікогенолітичну та гіперглікемічну дію катехоламінів. Тиреоїдні гормони беруть участь в регуляції виділення глюкокортикоїдів наднирниками, гормону росту аденогіпофізом.
У різних тварин доведена роль тиреоїдних гормонів в регуляції мінерального обміну, особливо в обміні кальцію і магнію. Після видалення щитоподібної залози у тварин в ранньому віці припиняється їх фізичний, статевий і психічний розвиток.
Гормони щитоподібної залози стимулюють окостеніння епіфізів трубчастих кісток, дозрівання хряща і таким чином прискорюють розвиток кісток. У цих процесах важливе значення має комбінована дія тиреоїдних гормонів і СТГ гіпофіза. Проте, останній регулює тільки ріст, а гормони щитоподібної залози – як ріст, так і диференціювання. Відмінності в швидкості росту тварин різних порід обумовлені різною інтенсивністю секреції йодовмісних гормонів щитоподібної залози.
Гормони щитоподібної залози впливають на ріст і розвиток шкіри та її похідних (волосся, пір’я), прискорюють скорочення серця. Вони беруть активну участь в регуляції рубцевих процесів у жуйних тварин. Під впливом гормонів посилюється бродіння кормової маси, зростає кількість ЛЖК, головним чином оцтової, прискорюється всмоктування їх, а також високомолекулярних жирних кислот і глюкози в кров, стають більш потужними і частішими скорочення передшлунків. Тиреоїдні гормони підвищують продукцію молока і вміст жиру в ньому. У великої рогатої худоби молочних порід щитоподібні залози функціонують активніше, ніж у м’ясних, у жирномолочних порід − інтенсивніше, ніж у корів з низьким вмістом жиру в молоці. Ін’єкція тироксину збільшує вміст молочного цукру – лактози в молоці.
Гормони щитоподібної залози впливають на розвиток нервової тканини. При природженій недостатності функції щитоподібної залози стає недорозвиненою центральна нервова система. При підвищенні функції щитоподібної залози в центральній нервовій системі збудливий процес переважає над гальмівним.
Фізіологічну гіпофункцію щитоподібної залози відзначають у тварин, що впадають у сплячку (ведмеді, їжаки, бабаки, кажани та ін.). Пробудження цих тварин пов’язано з підвищенням функції цієї залози. Введенням тироксину можна викликати передчасне пробудження їжаків, що знаходилися на зимівлі.
221
При патологічній зниженій функції (гіпофункції) щитоподібної залози волосся у тварин стає тьмяним, ламким і випадає, шкіра робиться шорсткою. При цьому припиняється розвиток статевих залоз. У дорослих тварин може відбуватися запліднення, але ембріони, як правило, гинуть в ранні терміни вагітності, оскільки тироксин необхідний для забезпечення імплантації заплідненої яйцеклітини в слизову оболонку матки. Підвищена функція (гіперфункція) щитоподібної залози також несприятливо позначається на діяльності статевих залоз: знижує їх чутливість до гонадотропних гормонів, що веде до порушення статевих циклів і переривання вагітності.
У високогірних областях, передгір’ях, болотистих, торф’яних і піщаних місцевостях, де в ґрунті є недостатня кількість йоду, можливе виникнення хвороби, що має назву «ендемічний зоб». При цьому захворюванні щитоподібна залоза, як правило, збільшується або в результаті розростання сполучної тканини (простий зоб), або внаслідок збільшення залозистої тканини (колоїдний зоб). Але в обох випадках кількість гормонів, що утворюються в ній, знижується порівняно з нормою.
Тиреокальцитонін − це гормон щитоподібної залози, що зберігає в організмі кальцій. Він знижує рівень іонів кальцію і фосфору в плазмі крові. Секреція цього гормону спостерігається тільки при значному збільшенні рівня кальцію і фосфору в крові. Дія гормону виявляється швидко і триває 2-3 год. Пониження кількості іонів кальцію в плазмі крові відбувається за рахунок ослаблення мобілізації його з кісток, а зниження вмісту фосфору – за рахунок посилення виділення фосфору з сечею. Тиреокальцитонін пригнічує функцію остеокластів, що руйнують кісткову тканину, і активізує функцію остеобластів, що формують її. Кальцитонін гальмує вихід органічної речовин з кісткового матриксу, як і мінеральних компонентів кістки. Рівень кальцитоніну підвищується у вагітних і лактуючих самок, і це свідчить про його фізіологічну роль в захисті організму матері від надмірних втрат кальцію. Кальцитонін гальмує секрецію гастрину, а гастрин стимулює секрецію кальцитоніну. Тиреокальцитонін діє на вміст кальцію в плазмі крові в протилежність гормону прищитоподібних залоз – паратгормону.
Регуляція функції щитоподібної залози. Діяльність щитоподібної залози регулюється центральною нервовою системою двома шляхами: по еферентних нервах і через гіпофіз. Щитоподібна залоза забезпечена розгалуженою мережею нервових волокон, які йдуть до неї з шийного симпатичного вузла, блукаючого, язикоглоткового і під’язикового нервів, а також тих нервових волокон, що надходять з нервових сплетень, розташованих поблизу загальної сонної і підключичної артерій.
Велику роль в регуляції функцій щитоподібної залози відіграють ретикулярна формація і гіпоталамус, що стимулюють її діяльність, посилаючи імпульси по симпатичних нервах. Крім того, гіпоталамус впливає на функцію щитоподібної залози через гіпоталамо-аденогіпофізарну систему, оскільки в його нейронах виробляється нейросекрет, що сприяє виділенню гіпофізом ТТГ. При зниженні секреції трийодтироніну і тетрайодтироніну вміст ТТГ гіпофіза в плазмі підвищується; і навпаки, концентрацію ТТГ можна знизити введенням
222
трийодтироніну і тетрайодтироніну. Вироблення ТТГ стимулюється нейросекретом (ТРГ) гіпоталамуса. Можливо також, що тиреоїдні гормони при накопичуванні в щитоподібній залозі здатні пригнічувати свій власний синтез і виведення Т4 в кров (аутокринний ефект).
Прищитоподібні (паращитоподібні) залози є в усіх класів хребетних тварин за винятком риб. У ссавців є одна або дві пари прищитоподібних залоз, що розташовані на поверхні щитоподібної залози або дещо попереду неї. Паренхіма цих залоз складається з двох видів клітин: головних і оксифільних. Секретуючими є головні клітини.
Після видалення прищитоподібних залоз (паратиреоектомії) підвищується збудливість нервової системи, тварини стають неспокійними, з’являються фібрилярні коливання окремих м’язів голови і тулуба. У подальшому коливання стають все сильнішими і переходять в напади загальних тетанічних судом. Особливо бурхливо судорожні припадки протікають у собак. Рослиноїдні тварини (за винятком кролів) легше переносять видалення прищитоподібних залоз. Це пояснюється наявністю у них додаткових прищитоподібних залоз, що зберігаються після операції, і великим вмістом кальцію в кормі. Повне видалення тканини прищитоподібих залоз смертельне для всіх видів тварин. Судоми, що виникають в результаті видалення цих залоз, мають центральне походження і пов’язані зі збудженням рухових центрів середнього і довгастого мозку внаслідок зниження концентрації іонів кальцію в плазмі крові. При цьому одночасно зростає вміст фосфору. Таким чином, основна функція прищитоподібних залоз – підтримка гомеостазу кальцію і фосфору.
У 1926 р. з прищитоподібних залоз бика був одержаний екстракт, який сприяв підвищенню рівня кальцію і зниженню концентрації фосфору в крові і підтримував життя тварин з видаленими прищитоподібними залозами. Згодом був отриманий гормон прищитоподібних залоз – паратгормон (ПТГ), що є білком з молекулярною масою близько 8500. За новою класифікацією ПТГ називають паратерином. Він посилює активність остеокластів – клітин, що руйнують кістки. При цьому іони кальцію вивільняються з кісткових депо і надходять в кров. Одночасно з кальцієм в кров виводиться і фосфор. Але під впливом паратерину різко посилюється виведення фосфатів з сечею. Тому концентрація фосфатів у плазмі крові знижується. Паратерин також підсилює всмоктування кальцію з кишечнику і реабсорбцію цього іона в дистальних ниркових канальцях, що сприяє підвищенню його концентрації в плазмі крові.
НАДНИРКОВІ ЗАЛОЗИ
Надниркові залози (наднирники) − парні органи, що розташовані медіокраніально відносно нирок. Вони складаються з двох частин, що різняться за своїми походженням, морфологією та функцією, − кіркової (кори) та мозкової речовин. Кіркову речовину ділять на клубочкову, пучкову і сітчасту зони. Кожна з цихзон містить характерні ферменти і виробляє специфічнігормони (рис. 9.6). Співвідношення ширини вказаних зон у кірковій речовині надниркової залози зрілого організму тварин становить відповідно 1:9:3. У птиці чіткого поділу
223
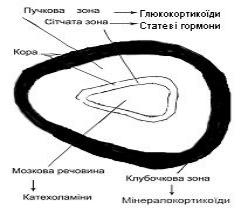
кіркового шару наднирників на зони немає; хромафінні клітини не мають чіткої локалізації в тканинах наднирників. Кожен наднирник оточений щільною капсулою. Надниркові залози інтенсивно постачаються кров’ю, артерії розгалужуються в капсулі або в кірковій речовині та утворюють анастомози.
Кіркова і мозкова речовини надниркових залоз мають різне походження. Їх можна розглядати як дві самостійні залози внутрішньої секреції. Кіркова речовина в ембріогенезі утворюється з целомічного епітелію, з якого також утворюються нирки і гонади; мозкова речовина – із клітинних елементів черевних симпатичних гангліїв. Ці особливості походження дозволяють зрозуміти тісний зв’язок гормонів кори наднирників зі статевими гормонами так само, як і аналогію в дії гормонів мозкової речовини і симпатичної нервової системи. Гормони надниркових залоз відіграють виключно важливу роль в регуляції обмінних процесів, підтриманні гомеостазу, рівня артеріального тиску і забезпечують захисні властивості організму та інші життєво важливі процеси.
Рис. 9.6. Схема секреторної будови наднирників
При видаленні наднирників життя тварин неможливе без підтримуючої гормональної терапії.
Кіркова речовина надниркових залоз. В
клітинах клубочкової зони кори надниркових залоз утворюються мінералокортикоїди (альдостерон, дезоксикортикостерон); пучкової зони − глюкокортикоїди (гідрокортизон, кортизон і кортикостерон); а в клітинах сітчастої зони − статеві гормони (андрогени, естрогени, прогестерон). Вважають, що справжніми гормонами є кортикостерон і гідрокортизон, які визначають усі функції наднирників. Решта речовин є продуктами обміну гормонів. Кортикостерон, як попередник альдостерону, частково теж утворюється в клубочковій зоні (рис. 9.6).
Мінералокортикоїди (альдостерон, дезоксикортикостерон) беруть участь у регуляції мінерального обміну (баланс електролітів). Найактивнішим мінералокортикоїдом є альдостерон. Секреція альдостерону призводить до затримки Nа+, виведенню К+ і Н+, збереженню об’єму циркулюючої рідини. Альдостерон затримує Nа+ і призводить до втрати К+ не тільки в нирках, але і в слинних залозах, дистальних відділах товстого кишечнику і потових залозах. Дезоксикортикостерон – проміжний продукт утворення альдостерону, володіє значно меншою (в 30 разів), ніж альдостерон, натрійзатримуючою дією та більш вираженим впливом на виділення кальцію.
Регуляція синтезу і секреції мінералокортикоїдів відбувається кількома шляхами. Перший механізм пов’язаний з дією ренін-ангіотензинової системи. Юкстагломерулярні клітини нирок у відповідь на зниження артеріального
224
тиску, об’єму крові або обох факторів разом починають секретувати ренін. Він впливає на ангіотензиноген, що надходить в кров з печінки і циркулює в ній у складі α2-фракції глобулінів, розщеплюючи його з утворенням ангіотензину I. Ця речовина перетворюється на ангіотензин II при проходженні крові переважно через легені. Ангіотензин ІІ стимулює клітини клубочкової зони наднирників, що секретують стероїдний гормон альдостерон. Крім того, він володіє судинозвужуючою дією на артерії і артеріоли, полегшує вивільнення норадреналіну, зменшує швидкість клубочкової фільтрації, збільшує реабсорбцію натрію, посилює почуття спраги, збільшує секрецію АДГ і АКТГ. Продукт метаболізму ангіотензину II − ангіотензин III, який теж є активним стимулятором секреції альдостерону. Відновлення артеріального тиску та об’єму крові, а також прямий інгібуючий ефект альдостерону й ангіотензину II за механізмом зворотного зв’язку призводять до ефективного гальмування секреції реніну.
Синтез і секрецію альдостерону клітинами клубочкової зони стимулюють не тільки ангіотензини II і III, але й АКТГ, простагландин Е, висока концентрація К+ і низька концентрація Nа+. До інгібіторів синтезу і секреції альдостерону відносяться дофамін, передсердний натрійуретичний фактор (ПНФ) і Nа+ у високій концентрації.
Глюкокортикоїди (гідрокортизон, кортизон і кортикостерон) безпосередньо або опосередковано регулюють практично всі фізіологічні і біохімічні процеси в організмі тварин. Їх дія проявляється на рівні генів в органах-мішенях шляхом вибіркової активації специфічних інформаційних і рибосомальної РНК, стимуляції синтезу білка з амінокислот, що доставлені транспортною РНК. Головним глюкокортикоїдом у мавп, овець і морських свинок є кортизон; у щурів, мишей, кроликів і птахів – кортикостерон, а у великої рогатої худоби, свиней, собак і кішок – обидва ці гормони.
Глюкокортикоїди впливають на обмін вуглеводів, білків і жирів, посилюють процеси утворення глюкози з білків (глюконеогенез), а також відкладення глікогену в печінці, є антагоністами інсуліну щодо регуляції вуглеводного обміну. Вони зумовлюють розпад тканинних білків, затримують включення амінокислот у білки організму шляхом пригнічення їх транспорту із плазми крові в клітини, а також прискорюють процес виділення азоту (катаболічний ефект). З іншого боку, гормони пучкової зони кори наднирників можуть викликати анаболічні реакції, такі, як індукція синтезу специфічних ферментів або навіть гіпертрофія органу (печінка). Прикладами анаболічних реакцій на глюкокортикоїди є стимуляція синтезу гормону росту в гіпофізі. Крім того, кортизол здатний змінювати реактивність клітин щодо інших гормонів та нейромедіаторів.
Глюкокортикоїди здатні виявляти протизапальну дію. Це пов’язано з тим, що названі гормони зменшують проникність стінки судин за рахунок зниження активності ферменту гіалуронідази, блокують секрецію серотоніну і гістаміну, кінінів і систему плазмін-фібринолізин. Протизапальна дія глюкокортикоїдів використовується в клінічній практиці. Крім того, ці гормони гальмують метаболізм у фібробластах, їх ділення та ріст. Найбільш яскравим проявом
225
пригнічення функції і ділення фібробластів під дією глюкокортикоїдів є уповільнення загоєння ран. Вони не тільки порушують утилізацію глюкози фібробластами, але й інгібують в них синтез ДНК, гальмуючи тим самим ділення клітин. Поєднання зниженої продукції колагену і сповільненого клітинного ділення істотно порушує репаративну фазу запального процесу.
Глюкокортикоїди беруть участь у багатьох процесах, пов’язаних з реакцією гіперчутливості, що зумовлена дією антитіл з екзогенними або ендогенними антигенами.
У регуляції функції глюкокортикоїдів бере участь гіпоталамус. В нейросекреторних клітинах ядер гіпоталамуса виробляється рилізинг-гормон (кортиколіберин), що надходить через ворітну систему в аденогіпофіз і сприяє там утворенню й вивільненню кортикотропіну. Вплив останнього на процес утворення глюкокортикоїдів здійснюється за принципом прямого і зворотного зв’язку. Кортикотропін стимулює секрецію глюкокортикоїдів у надниркових залозах, а надлишок їх у крові призводить до гальмування синтезу кортикотропіну в аденогіпофізі. Таким чином, гіпоталамус, аденогіпофіз і кора надниркових залоз перебувають в тісному функціональному зв’язку, утворюючи єдину гіпоталамо- гіпофізарно-надниркову систему.
Глюкокортикоїди разом з іншими гормонами (АКТГ) сприяють адаптації організму до нових умов існування, а також до дії різних несприятливих чинників: зміна температури навколишнього середовища, кормовий стрес, кисневе голодування і інше. Тому їх називають захисними чи адаптаційними гормонами.
При дії різноманітних патогенних і надмірних за силою й тривалістю подразників (інтоксикація, інфекція, надмірне фізичне навантаження, переохолодження, гіпоглікемічні стани тощо) на організм тварин виникає неспецифічна захисна, пристосувальна реакція або комплекс реакцій, який називається загальним адаптаційним синдромом. Фактори, що викликають цю неспецифічну реакцію, прийнято називати стресорами, оскільки загальний адаптаційний синдром виникає при такому стані організму, який канадський вчений Ганс Сельє назвав станом «напруження» (stress). При стресі збуджується симпатична нервова система і з мозкового шару наднирників у кров надходять адреналін і норадреналін. Під їх впливом у гіпоталамусі збільшується утворення кортиколіберину, що сприяє підвищеній секреції в передній частині гіпофіза АКТГ і глюкокортикоїдів в пучковій зоні наднирників.
Статеві гормони сітчастої зони кори наднирників. У сітчастій зоні продукуються статеві гормони – аналоги стероїдних гормонів, що виробляються в статевих залозах. Ці гормони мають значення для розвитку статевих органів у ранньому постнатальному віці і появи вторинних статевих ознак у той період, коли внутрішньосекреторна функція статевих залоз ще незначна. Вони (особливо андрогени) сприяють обміну білків, стимулюючи синтез їх в організмі, впливають на затримку натрію, води, фосфору і кальцію, сприяють кальцифікації кісток, а при надмірному утворенні викликають гіпертрофію сальнихзалоз.
226
До андрогенів відносяться андростендіон, 1-оксіандростендіон і дегідроепіандростендіон. Найбільша гормональна активність властива андростендіону, проте вона в п’ять разів нижча, ніж у тестостерону, що утворюється в сім’яниках. Наявність андрогенів в наднирниках не залежить від статі тварини і має особливе значення в період достатевого дозрівання, забезпечуючи анаболічну дію, посилюючи ріст тіла і розвиток скелетної мускулатури. Естрогени, в основному естрон і естрадіол, виробляються в сітчастій зоні наднирників також в дуже малій кількості. Їх вплив проявляється у випадках розвитку пухлини кори наднирників. Прогестерон утворюється в корі наднирників з холестерину в процесі утворення інших стероїдних гормонів, а під час подальших перетворень може бути попередником 11дезоксикортикостерону й альдостерону. Його дія на матку та інші прогестерончутливі тканини аналогічна гормону жовтого тіла.
Регуляція секреції статевих стероїдів, що виробляються в сітчастій зоні наднирників, здійснюється за допомогою тропних гормонів аденогіпофіза: ФСГ для естрогену і ЛГ для андрогенів і прогестерону. Разом з цим, ріст пучкової і сітчастої зон, а також утворення і секреція глюкокортикоїдів стимулюються адренокортикотропним гормоном (АКТГ). АКТГ стимулює і утворення дегідроепіандростерону і андростендиолу у зародків ссавців, тоді як у дорослих тварин основним регулятором секреції андрогенів наднирників стає ЛГ.
Мозкова речовина надниркових залоз. В ембріогенезі мозкова речовина наднирників і симпатична нервова система розвиваються з одного зачатку. Секреторні клітини мозкової речовини забарвлюються фарбами, що містять солі хрому, в коричневий колір і тому називаються хромафінними. Різні клітини відрізняються забарвленням: клітини, що мають темне забарвлення, виробляють норадреналін, світліші клітини − адреналін. Ці клітини являють собою видозмінені симпатобласти та іннервуються прегангліонарними нервовими волокнами симпатичної нервової системи. Деякі з них диференціюються в нейрони.
У хромафінних клітинах виробляються катехоламіни (адреналін, норадреналін, дофамін), що утворюються з амінокислоти фенілаланіну. Фенілаланін під впливом ряду ферментів перетворюється в тирозин, деоксифенілаланін, дофамін і, нарешті, в норадреналін. Завдяки наявності в мозковій речовині наднирників ферменту фенілетаноламін-метилтрансферази норадреналін перетворюється в адреналін. Далі адреналін запасається для секреції в секреторних гранулах. При злитті мембрани гранул з плазматичною мембраною клітини гормон вивільнюється.
Вміст і співвідношення різних форм катехоламінів у мозковій речовині наднирників неоднаковий у різних видів тварин. Так, у наднирниках кролів і морських свинок адреналін становить 95 %, у худоби ~ 80, у котів ~ 50 %. У домашньої птиці переважає норадреналін, досягаючи 80 % всіх катехоламінів. У щурів рівень секреції адреналіну і норадреналіну складає 1:1.
За аналогією з глюкокортикоїдами більша частина адреналіну і норадреналіну в крові зв’язана з альбумінами, а менша – у вільному активному стані.
227
Катехоламіни володіють широким спектром дії на організм. Їх фізіологічна дія відрізняється як у кількісному, так і в якісному відношенні. Ці гормони здійснюють значний вплив на мозок, серцево-судинну систему, скелетну мускулатуру, гладеньку мускулатуру травного каналу, матки, очей і бронхів, центральну нервову систему, селезінку і на процес зсідання крові. За дії катехоламінів підвищується хвилинний об’єм серця, частота пульсу, артеріальний тиск. Звуження судин внутрішніх органів і розширення судин скелетної мускулатури зумовлює перерозподіл кровотоку. Окрім того, скорочується селезінка, що призводить до збільшення об’єму циркулюючої крові і кількості в ній еритроцитів.
Адреналін впливає на вуглеводний обмін, посилює розпад глікогену, зумовлюючи зменшення його запасів у печінці та м’язах, тобто є антагоністом інсуліну. Підвищуючи вміст глюкози в крові цей гормон активує інкрецію інсуліну, в той же час безпосередньо пригнічує функцію панкреатичних острівців. Адреналін може значно збільшувати працездатність скелетних м’язів. При цьому він прискорює глікогеноліз з накопиченням лактату, який частково переноситься в печінку, де перетворюється на глікоген. Потім глікоген розпадається до глюкози і знов надходить в кровотік. Лактат потрапляє в серце, де ефективно використовується як джерело енергії. Під впливом адреналіну через α-адренорецептори зменшується прояв явищ втоми в скелетних м’язах, підвищується збудливість рецепторів зорового, слухового та вестибулярного аналізаторів тощо, завдяки чому поліпшується сприйняття організмом зовнішніх стимулів.
Адреналін чинить ліполітичну дію – підвищує вміст вільних жирних кислот у крові. За рахунок цього різко збільшується використання кисню тканинами, оскільки значно підвищується окислення жирних кислот і зростає теплоутворення. При фізичних навантаженнях підвищується виділення адреналіну, прискорюється ліполіз і працюючі м’язи забезпечуються енергетичним матеріалом – жирними кислотами. Під впливом адреналіну посилюється енергетичний обмін, у тому числі й основний.
Адреналін спричиняє прискорення і посилення серцевих скорочень, поліпшує проведення збудження в серці, звужує через α1-адренорецептори артеріоли шкіри, органів черевної порожнини, підвищуючи таким чином артеріальний тиск. Адреналін через β-адренорецептори пригнічує скорочення гладеньких м’язів шлунка та кишок, зумовлює при подразненні β- адренорецепторів послаблення бронхіальних м’язів, внаслідок чого просвіт бронхів і бронхіол розширюється. Водночас, адреналін спричиняє скорочення радіальних м’язів райдужної оболонки ока, внаслідок чого зіниці розширюються. Під впливом адреналіну через α-адренорецептори також скорочуються піломотори шкіри, що призводить до піднімання волосся.
Норадреналін має ознаки гормону та медіатору (трансмітера), оскільки виконує функції передавача збудження синаптичних нервових закінчень на ефектор, а також у нейронах ЦНС. На відміну від адреналіну, норадреналін має переважно загальну судинозвужуючу дію на периферійні судини (шкіри, кінцівок, органів черевної порожнини, м’язів, мозку), що проявляється
228
підвищенням як максимального, так і мінімального артеріального тиску; істотно не прискорює ритм серця, не змінює систолічний об’єм серця, покращує коронарний кровотік, не впливає на тонус бронхів, матки, кишок, жовчного міхура, володіє слабко вираженою глікогенолітичною дією, не збуджує центральну нервову систему.
Дія дофаміну, в основному проявляється підвищенням моторної активності, ниркового кровотоку, позитивним інотропним та хронотропним впливом на серце.
Надходження в кров адреналіну і норадреналіну супроводжується підвищенням збудження симпатичної нервової системи. Ці катехоламіни посилюють і подовжують вегетативні ефекти. Тому прийнято говорити про симпатико-адреналову систему. Отже, катехоламіни відіграють важливу роль у пристосувальних захисних реакціях організму, що зумовлюють екстренну перебудову функцій, спрямованих на підвищення адаптації організму до надзвичайних умов внутрішнього та навколишнього середовища.
Гормони мозкової речовини наднирників не накопичуються в організмі і володіють короткочасною дією. Катехоламіни після надходження у кровоток швидко руйнуються. Частина адреналіну, норадреналіну і продуктів їх перетворення зв’язується в печінці з глюкуроновою і сірчаною кислотами й виводиться з організму з сечею.
ЕНДОКРИННА ФУНКЦІЯ ПІДШЛУНКОВОЇ ЗАЛОЗИ
Підшлункова залоза – це орган, який складається з екзокринної та ендокринної частин. В екзокринній частині виробляється травний сік, що надходить в дванадцятипалу кишку і містить необхідні для розщеплення білків, ліпідів і вуглеводів ферменти. В ендокринній частині – панкреатичних острівцях Лангерганса – синтезуються гормони, що регулюють метаболічні процеси, особливо обмін вуглеводів.
Упанкреатичних острівцях залозистими клітинами виробляються два основні гормони підшлункової залози: інсулін і глюкагон. Клітини, що синтезують інсулін, − базофільні, їх називають β-(В-)клітинами; клітини, що виробляють глюкагон, – ацидофільні α-(А-)клітини. Існує третій тип клітин – дефінитивні δ-(Д-) клітини, в яких синтезується соматостатин. Окрім того, у Д1- клітинах синтезується вазоактивний інтерстиціальний поліпептид (ВІП) і гастрин, у РР-клітинах – панкреатичний поліпептид. З екстрактів інкреторної тканини підшлункової залози виділені також ваготонін, центропнеїн, ліпокаїн, бомбезин.
Інсулін − загальний цукрозберігаючий гормон організму. Молекула інсуліну складається з двох амінокислотних ланцюгів – А і В, сполучених двома дисульфідними містками. Цинк, що входить до складу гормону, забезпечує його депонування в гранулах β-клітин.
Укрові інсулін міститься в двох формах: вільній і зв’язаній з білками. Зв’язаний інсулін є відповідним резервом, що може використовуватися під час надходження в кров великої кількості глюкози. Інсулін, який не брав участь в процесах обміну речовин, руйнується ферментом інсуліназою.
229
Інсулін є анаболічним гормоном з широким спектром дії. Його роль полягає в підвищенні синтезу вуглеводів, жирів, білків. Він стимулює метаболізм глюкози.
Інсулін впливає на проникність глюкози у мембрани клітин міокарду та скелетних м’язів. Для надходження глюкози в клітини головного мозку інсулін не потрібен. Але він необхідний для проходження амінокислот крізь гематоенцефалічний бар’єр. Інсулін стимулює синтез глікогену в печінці, уповільнює процес глюконеогенезу (утворення глюкози з амінокислот), впливає на обмін ліпідів, підсилюючи здатність жирової тканини і печінки до накопичення їх у формі тригліцеридів. Стимулюючи синтез білків і глікогену, гормон впливає на розвиток, диференціацію і трансформацію клітин, зокрема активує Т- і В-лімфоцити. Інсулін стимулює ріст та проліферацію клітин, впливає на синтез соматомедину в печінці і, можливо, в інших тканинах. Панкреатична β-клітина акумулює інсулін в секреторних гранулах у готовій для секреції формі.
Зміна рівня глюкози в крові сбереться спеціалізованими клітинами – глюкорецепторами підшлункової залози, каротидного синусу, паравентрикулярних ядер гіпоталамуса. Збільшення вмісту глюкози в крові спричинює секрецію інсуліну. Швидкість виділення інсуліну прямо пропорційна вмісту глюкози в крові: вона тим вище, чим вище рівень глюкози.
Глюкоза та її метаболіти, маноза, лейцин – активні стимулятори синтезу проінсуліну і секреції інсуліну. Таким чином, в нормі процеси синтезу і секреції гормону, як правило, взаємопов’язані, але регулюються різними внутрішньоклітинними сигналами. Іншими стимуляторами синтезу інсуліну є гормон росту та глюкагон, останній з яких здійснює свій регулюючий вплив тільки в присутності глюкози. В той же час синтез проінсуліну інгібується адреналіном.
Постійний рівень синтезу проінсуліну може зберігатись досить довго (дні, а навіть і тижні). Це пов’язано зі збільшенням або зменшенням кількості β- клітин. Синтез проінсуліну значно уповільнюється при голодуванні або низькому вмісті вуглеводів і високому вмісті жирів у кормі. Підвищення його синтезу відбувається при згодовуванні корму з високим вмістом вуглеводів, при ожирінні, вагітності та в умовах хронічного надлишку гормону росту.
Антагоністом інсуліну є другий гормон підшлункової залози, що виробляється в α-клітинах, − глюкагон. Глюкагон є пептидом, що складається з 29 амінокислот. Амінокислотна послідовність глюкагону більшості видів ссавців однакова. Визначено два типу глюкагону: панкреатичний і кишковий (ентероглюкагон).
Подібно до інсуліну, глюкагон утворюється з його попередника. При стимуляції секреторної активності β-клітин гормон виділяється в кров шляхом екзоцитозу. Глюкоза і Са2+, стимулюючи секрецію інсуліну, інгібують секрецію глюкагону. Причому, для секреції гормонів підшлункової залози як у β-, так і в α-клітинах необхідна глюкоза в окисненій формі.
Глюкагон впливає головним чином на печінку, де негайно стимулює глікогеноліз, а через більш тривалий час − глюконеогенез і кетогенез.
230