

Физиология Karpovskyi
.pdfхімусу з утворенням кінцевих продуктів гідролізу білків, жирів та вуглеводів. А ось під дією гнилісних бактерій утворюються отруйні для організму сполуки: крезол, фенол, індол, скатол, путресцин, кадаверин, NH3 та інші, що всмоктуються в кров та знешкоджуються у печінці, утворюючи парні сполуки з сульфатною (сірчаною) та глюкуроновою кислотами, а з NH3 в гепатоцитах в орнітиновому циклі синтезується сечовина. Жовчний пігмент білірубін перетворюється на стеркобілін. Разом з тим слід відзначити, що мікрофлора товстих кишок має важливе значення у життєдіяльності організму й визначається такими функціями:
-Утворенням імунологічного бар’єру та антагонізму по відношенню до патогенних мікробів. Тому дуже тяжко протікають дисбактеріози, що обумовлені порушенням екосистеми товстих кишок і виявленням мікроорганізмів, які не властиві мікробному обсіменінню в товстих кишках здорового організму.
-Кишкова мікрофлора здійснює гідроліз багатьох органічних речовин, синтезує вітамін К, деякі вітаміни групи В, а також інші біологічно активні сполуки.
-Для процесів травлення у товстих кишках важливе значення мають бактерії, що розщеплюють клітковину. Якщо у жуйних тварин целюлозолітичні бактерії заселяють переважно передшлунки, то у коня клітковина гідролізується в основному у сліпій кишці, що є своєрідним “другим шлунком , який виконує функції рубця. У сліпій кишці коня перетравлюється до 50% клітковини, до 40% білка, відбуваються бродильні процеси з утворенням великої кількості газів – СО2, NH3, SH, CH4 та інших. Травні процеси мають місце і у великій ободовій кишці, у той час як у малій ободовій
–їх майже немає.
Утовстих кишках свиней вміст перебуває у межах 16–20 год. Виявлено, що у цей відділ травного тракту надходять до 14% вуглеводів та 12% білка корму, а перетравлюється лише до 9% вуглеводів та до 3% білка. Отже, питома вага у травленні поживних речовин корму товстих кишок свиней не має суттєвого значення.
Ужуйних тварин у товстих кишках перетравлюється та усмоктується у кров до 15–20% клітковини корму.
Товсті кишки відіграють важливу екскреторну роль. У їх просвіт надходить багато кінцевих продуктів обміну, мінеральних речовин, а у каудальних ділянках відбувається згущення вмісту за рахунок резорбції води та формування фекалій.
Рухова діяльність товстих кишок вивчається тими ж методами, що й тонких. Основними видами руху товстих кишок є маятникоподібні, що сприяють перемішуванню вмісту, а також перистальтичні та антиперистальтичні скорочення.
Для гладеньких м’язів товстих кишок характерна ще й автоматія, хоча менш виражена, ніж у тонких кишках.
Регуляція секреторної та моторної функцій товстих кишок здійснюється за такими ж фізіологічними механізмами, що й тонких кишок.
РЕГУЛЯЦІЯ МОТОРИКИ КИШОК
Міогенні механізми регуляції. Гладеньком’язові клітини кишки володіють здатністю спонтанно скорочуватися, що забезпечується міогенним механізмом. Міогенним механізмом регуляції моторики тонких кишок пояснюється також скоротлива реакція гладеньком’язових клітин під час перистальтики.
Не виключено, що у механізмі міогенної регуляції моторики кишок беруть участь також нервові механізми (тензорецептори, пропріорецептори та інші), м’язові оболонки та інтрамуральна нервова системі, що являє собою міоентеральне (ауербахове) глибоке м’язове та підслизове (мейснерове) сплетення, що замикають місцеві рефлекторні дуги. Механічне подразнення кишки викликає скорочення у цій самій ділянці та просування її вмісту у аборальному напрямку.
Нервова регуляція моторики кишок. Нервові центри, що регулюють моторну діяльність кишок, розміщені у довгастому мозку, а вищі центри – у гіпоталамусі та корі великих півкуль головного мозку. Еферентними нервами є блукаючі та черевні, вони передають збудливі та гальмівні впливи на моторику кишок. У складі цих нервів містяться збудливі та гальмівні волокна. Симпатичні волокна переважно гальмують, а парасимпатичні переважно збуджують моторну діяльність травного тракту.
У механізмі нервової регуляції моторної діяльності кишок велике значення має рефлекторна регуляція. Так, подразнення рецепторів шлунка рефлекторно збуджує моторну діяльність кишок. Зараз до моторних рефлексів кишок включають: 1) стравохідно-кишковий; 2) шлунково-кишковий; 3) кишково-кишковий. Установлено, що під час подразнення інтерорецепторів стравоходу рефлекторно посилюється моторика кишок. Тоді при збудженні імпульси надходять до гладенької мускулатури кишок блукаючими, а гальмівні
– черевними нервами.
Слід відзначити, що під час сильного (неадекватного) подразнення будьякої ділянки кишки виникає гальмування моторної діяльності як вищерозміщених, так і нижчерозташованих щодо неї частин.
Рефлекторне гальмування моторної діяльності кишок є можливим за умов подразнення інших систем організму: сечоводів, сечового міхура, ниркових мисок, очеревини, матки, дихальної та інших систем організму та звичайно, структур головного мозку, включаючи кору великих півкуль. Окрім того, можливі умовно-рефлекторні зміни моторної функції кишок, як збуджуючі, так і гальмівні впливи.
УСМОКТУВАННЯ
Усмоктуванням називається процес проникнення різних речовин у кров або лімфу через біологічні мембрани. До останніх належать епітеліальні шари шкірних покривів, слизові оболонки травної системи, жовчного міхура, легеневих альвеол, ендотелію, що вистеляє серозні оболонки черевної порожнини, суглобових сумок, ендотеліальний шар капілярів, епітелій сечових канальців нирки та інше. Таким чином, усмоктування може здійснюватися через усі тканини організму. Проте, морфологічно та функціонально до цього
процесу пристосовані клітини слизової оболонки травної системи, особливо тонкого відділу кишок.
Усі біологічні мембрани є напівпроникливими, тобто прохідними для одних речовин та непрохідними для інших. Взагалі клітинні мембрани проникливі для речовин, що утворюють іоннота молекулярно-дисперсні речовини й непроникливі для речовин, які перебувають у колоїдному стані.
МЕХАНІЗМ УСМОКТУВАННЯ
У механізмі усмоктування важлива роль належить клітинним мембранам, їх проникливості та властивостям. На процес усмоктування впливає ряд фізичних явищ: дифузія, осмос, фільтрація, тиск у порожнині та інше. Під час процесу усмоктування речовина минає мембрану клітини, мікроканальці ендоплазматичної сітки, базальну мембрану, сполучну тканину та ендотелій капілярів.
Деякі речовини можуть проходити через міжклітинну щілину.
За принципом осмосу та дифузії можуть надходити до крові О2, СО2, NH3, Н2О. Цими фізико-хімічними закономірностями не можна пояснити усмоктування продуктів гідролізу органічних сполук. Багато з речовин переходять з кишок до крові супроти концентраційного градієнта, тобто надходять від малої концентрації до великої. Такий процес усмоктування супроводжується витратами енергії макроергічних сполук. Окрім того має місце вибіркове усмоктування деяких метаболітів, а також транспорт різних речовин за участі специфічних білків-переносників та інше.
УСМОКТУВАННЯ У РІЗНИХ ДІЛЯНКАХ ТРАВНОГО ТРАКТУ
Уротовій порожнині усмоктуються як правило легкорозчинні речовини, котрі легко проникають через мембрани смакових цибулин та обумовлюють відчуття смаку корму, що береться. З ротової порожнини усмоктуються до крові деякі лікарські речовини та деякі продукти гідролізу органічних сполук. Проте у зв’язку з короткочасним перебуванням корму у ротовій порожнині поживні речовини не встигають гідролізуватися (рис. 22).
Ушлунку можуть усмоктуватися багато які поживні речовини – амінокислоти, моносахариди, гліцерол, спирти, мінеральні сполуки. Проте, об’єм речовин, що всмоктуються, не перевищує кількості утвореного шлункового соку.
Упередшлунках жуйних тварин усмоктуються продукти перетравлювання та зброджування поживних речовин корму, незважаючи на те, що слизова оболонка передшлунків устелена багатошаровим плоским епітелієм. Багаточисельні сосочки рубця, сіточки та складки, значно збільшують резорбційну поверхню, а епітеліальні клітини чисельно васкуляризовані, містять багаточисельні ферменти та сульфгідрильні групи.
Із передшлунків легко усмоктуються до крові низькомолекулярні карбонові кислоти, NH3, амінокислоти, СО2 та мінеральні речовини. З книжки усмоктується близько 70% летких жирних кислот, що утворюються в рубці та до 100 літрів Н2О за добу.
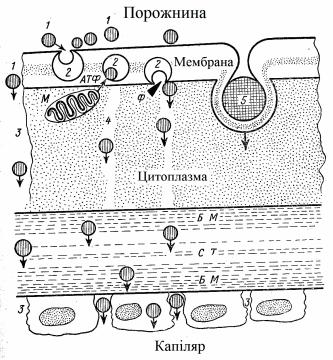
Основним місцем резорбції поживних речовин корму є тонкі кишки, що обумовлене морфофункціональними особливостями їх слизової оболонки та тим, що у цьому відділі проходить основний гідроліз білків, жирів та вуглеводів.
Слизова оболонка тонких кишок побудована з ворсинок, що мають значну васкуляризацію, лімфатичну систему, гладенькі м’язові волокна з іннервацією підслизового шару. Ворсинки покриті шаром епітеліальних клітин з багаточисельними виростами (мікроворсинками) до 3000 на кожній клітині, а на 1 мм2 поверхні кишкового епітелію – від 50 до 200 млн. мікроворсинок. Завдяки наявності мікроворсинок поверхня
усмоктування кишки збільшується у 14-39 разів.
Усмоктування білків. У тонких кишках усмоктуються продукти гідролізу білків – амінокислоти, а також небілкові азотисті сполуки: амоніак, пуринові та піримідинові основи, а також окремі мононуклеотиди. Інтенсивність усмоктування амінокислот залежить від співвідношення між циклічними та ациклічними, замінними та незамінними амінокислотами, а також функціонального стану війчастого апарату слизової оболонки кишки.
У механізмі всмоктування амінокислот є місце для активного переносу їх через стінку кишки, про що свідчить високий ступінь акумуляції амінокислот проти градієнту концентрації, а також гальмування цього процесу цианідом, 2, 4-динітрофенолом та іншими інгібіторами синтезу АТФ.
На рівні з активним транспортом амінокислот існує резорбція амінокислот за допомогою носія. Передбачають існування декількох “транспортних груп з особливим носієм для кожної групи: 1) нейтральні амінокислоти; 2) двоосновні амінокислоти та цистин; 3) кислі амінокислоти. За таких умов багато які амінокислоти мають два або навіть більше носіїв.
Показано, що у овець та телят відзначається переважно позитивна артеріовенозна (за участі ворітної вени) різниця за альбуміном та β-глобуліном сироватки. Останні поглиналися із крові, що притікає до травної системи овець та телят та використовувалися після гідролізу для підтримання амінокислотного гомеостазу у вмісті кишок. Під час утримання тварин на раціонах з недостатньою кількістю перетравного протеїну підвищувалася інтенсивність поглинання із крові ендогенних білків стінкою шлунковокишкового тракту жуйних тварин. Вивчаючи обмін нітрогеновмісних речовин між кров’ю та травною системою у жуйних тварин, виявлено позитивну А-В
різницю за сечовиною. Остання дифундує до порожнини шлунково-кишкового тракту, гідролізується під дією ферменту уреази (карбамідамідогідролази) до СО2 та NH3 з подальшим використовуванням нітрогену NH3 для утворення амінокислот. Процес фіксації NH3 практично у всіх тканинах відбувається шляхом відновлювального амінування α-кетоглутарату з утворенням глутамінової кислоти.
Усмоктування вуглеводів у кишках відбувається у формі моносахаридів. Установлено, що швидкість резорбції окремих моносахаридів залежить від
швидкості їх фосфорилювання, яке спливає за схемою:
глюкоза + АТФ гексокіназа→глюкозо-6-фосфат + АДФ Швидше за усіх усмоктується галактоза, потім глюкоза, далі – фруктоза
та дуже повільно усмоктується маноза.
Для забезпечення усмоктування глюкози проти градієнту концентрації потрібен натрій. Виявилося, що транспорт цукру та натрію взаємозалежний. Не тільки натрій стимулює транспорт цукру, а й цукор у свою чергу збільшує потік натрію.
У кишках жуйних тварин усмоктується невелика кількість глюкози, оскільки значна частина вуглеводів корму вже у передшлунках перетравлюється, зброджується до летких карбонових кислот і у такому стані усмоктується.
Усмоктування ліпідів. Відомо, що ліпіди та продукти їх гідролізу, погано або зовсім не розчиняється у воді (за винятком гліцерину), тому вони повинні зазнавати певних фізико-хімічних (дрібненя частинок та їх емульгація) та біохімічних (гідроліз) перетворень для переходу у водне середовище. Під час процесу гідролізу ліпідів у кишках створюються умови для трансформації нерозчинних у воді жирів та продуктів їх гідролізу до водорозчинних фізикохімічних форм. Відомо, що панкреатична ліпаза діє тільки на складні жири, що емульговані у воді. Основним емульгатором у тонких кишках є жовч, яка знижує поверхневий натяг у системі жир-вода, сприяє емульгації жирів, створює умови для прояву ліполітичної дії панкреатичної ліпази. Установлено, що під впливом панкреатичної ліпази від тригліцеридів від єднуються жирні кислоти, що знаходяться у крайніх положеннях молекули тригліцериду. Тому основним продуктом панкреатичного ліполізу тригліцеридів є 2-моногліцерид. За даними О.С. Фрезера жири корму усмоктуються у формі емульсії. Єдиною ефективною емульгуючою системою є система, що складається з таких компонентів: солей жовчних кислот, вільних жирних кислот та моногліцеридів. Таким чином, під дією панкреатичної ліпази, емульговані жири поступово переходять у формі моногліцеридів та жирних кислот до міцелярного стану. Потім жовчні кислоти забезпечують транспорт жирних кислих та моногліцеридів у формі міцели до слизової оболонки, де вони усмоктуються шляхом пасивної дифузії.
Отже, солі жовчних кислот беруть активну участь в усіх етапах травлення та всмоктування жиру: емульгування, гідролітичне розщеплення, активування ліпази, солюбілізація продуктів гідролізу та транспортування жирних кислот та моногліцеридів до епітеліоцита слизової оболонки кишки.
Встановлено, що у епітеліоцитах відбувається ресинтез тригліцеридів. Саме цим пояснюється наявність у лімфі ліпідів переважно у формі тригліцеридів.
Основні етапи біосинтезу тригліцеридів у клітинах слизової оболонки кишок:
1)активація жирних кислот ферментом тіокіназою у присутності АТФ та коферменту А (КоА) за реакцією:
жирна кислота + HS-КоА + АТФ → ацил-КоА + АМФ + пірофосфат (ФФ)
2)активовані жирні кислоти реагують з α-гліцерофосфатом з утворенням фосфатидної кислоти;
3)дефосфорилювання фосфатидної кислоти за допомогою ферменту фосфатази та утворення дигліцериду;
4)ацилювання дигліцериду до тригліцериду за реакцією
ацил-КоА + дигліцерид тригліцерид + HS-КоА
Окрім такого механізму можливий реосинтез тригліцеридів у слизовій оболонці кишки шляхом прямої етерифікації моногліцеридів активованими жирними кислотами. За сучасними уявленнями при усмоктуванні жирів велике значення має довжина ланцюга жирних кислот. Жирні кислоти з довгим ланцюгом від С16 та вище усмоктуються виключно до лімфи у формі тригліцеридів, що входять до складу хіломікронів, у той час як жирні кислоти з коротким ланцюгом достатньо швидко усмоктуються до системи ворітної вени, де виявляються у вільному стані.
Усмоктування холестеролу. У порожнині тонких кишок холестерол перебуває переважно у вільному стані (85–90% від загального вмісту холестеролу) і лише 10–15% – у формі стеридів. Розщеплення стеридів відбувається під дією холестерол-естерази панкреатичного та кишкового соків. Вільний холестерол бере участь разом з жирними кислотами, солями жовчних кислот та продуктами гідролізу тригліцеридів у формуванні міцелярної фази хімусу, що потрібна для усмоктування холестеролу.
Встановлено, що вільний холестерол уже в слизовій оболонці кишки має можливість реестерифікуватися, причому, переважно разом з ненасиченими довголанцюговими жирними кислотами. Окрім того у слизовій оболонці кишки відбувається синтез значної кількості холестеролу. На долю печінки і тонкої кишки випадає до 90% холестеролу, що синтезується у організмі.
Усмоктування води та мінеральних сполук. Усмоктування води та мінеральних речовин відбувається з поверхні слизової оболонки усього шлунково-кишкового тракту. Високий вміст води у травному тракті пов’язаний з надходженням її у складі травних соків. Так, установлено, що у корів кишками за добу переміщується до 150–180 л. води, з них більше ніж 100 л. – це вода травних соків. Окрім того, за умов високої концентрації у кишках солей, що погано усмоктуються, вода може переходити з крові в просвіт кишки. На цьому базується проносна дія деяких солей (Na2SO4, MgSO4).
Основним місцем усмоктування води є кишки. За даними О.В. Квасницького у свині за добу усмоктується у кишках до 21 л. води (біля 1 л. на 1 м кишки), а у товстих кишках – до 2 л. (біля 0,5 л. на 1 м кишки). У корів
тонкими кишками за добу проходить від 15 до 180 л. води і тільки 10% її виділяється з фекаліями.
У механізмі усмоктування води велике значення мають закономірності осмосу, а також мінеральні солі. Дослідження показують, що транспорт води дуже тісно сполучений із транспортом солей і, передусім Na+ та Cl–.
Слід зазначити, що усмоктування мінеральних речовин відбувається у вигляді неорганічних та органічних сполук. За цих умов відзначається вибірковість усмоктування, залежність його від розчинності солей та ступеня їх дисоціації.
Із катіонів найбільш швидко усмоктується К+, потім – Na+, повільніше – Са2+ та Mg2+, а з аніонів швидше усмоктується Clˉ та повільніше – сульфати та фосфати. Має значення й концентрація розчинів. Швидше усмоктуються мінеральні речовини з гіпотонічних та ізотонічних розчинів. Окрім того, слід мати на увазі, що є конкурентна взаємодія механізмів транспорту не тільки для різних іонів, а й навіть для одного катіона або аніона. За таких умов виділяють транспорт іонів за концентрованим градієнтом, активним транспортом, за участю специфічних переносників з утворенням різних комплексних сполук.
РЕГУЛЯЦІЯ УСМОКТУВАННЯ
Усмоктуванням завершуються травні процеси. Продукти гідролізу органічних сполук, вітаміни, вода, мінеральні речовини, а також продукти, що утворюються у шлунково-кишковому тракті, усмоктуються до крові та до лімфи. Інтенсивність цього процесу залежить, передусім, від проникливості кишкових структур шлунково-кишкового тракту, кровоносних та лімфатичних капілярів та багатьох інших факторів. Усмоктування перебуває у тісному взаємозв’язку з іншими функціями травної системи – секреторною, моторною та метаболічною.
Так само, як і усі інші функції організму, усмоктування у організмі перебуває під контролем центральної нервової системи та гуморальних факторів регуляції. Нервові центри, що контролюють процес усмоктування розміщені у довгастому мозку, гіпоталамусі та у корі великих півкуль, оскільки є можливими умовно-рефлекторні впливи на усмоктування речовини, що надходять із шлунково-кишкового тракту до крові та лімфи.
Еферентні нервові імпульси надходять до різних морфологічних структур, що причетні до усмоктування, симпатичними та парасимпатичними нервовими волокнами. Як правило, парасимпатичними нервовими волокнами надходять сигнали, що стимулюють процес усмоктування, який пов’язаний як із підвищенням проникності клітинних мембран, секреторною та моторною діяльністю травного апарата, так із утворенням специфічних білківпереносників, а симпатичними нервовими волокнами надходять нервові імпульси, що знижують інтенсивність резорбції.
На процес усмоктування суттєвий вплив мають гормони. Так, гормон підшлункової залози інсулін стимулює усмоктування глюкози, вільних амінокислот. Підвищує усмоктування глюкози та летких жирних кислот гормон
тироксин, тоді як пітуїтрин зменшує резорбцію глюкози. Адреналектомія викликає зниження усмоктування продуктів гідролізу вуглеводів та жирів. Посилює усмоктування амінокислот соматотропний гормон гіпофіза.
Стимулює усмоктування кальцію та фосфору вітамін Д3. Жовчні кислоти сприяють усмоктуванню жирних кислот та холестеролу. На швидкість усмоктування також впливає співвідношення між поживними речовинами, продуктами гідролізу органічних сполук, мінеральними речовинами та енергозабезпеченістю організму, оскільки усмоктування більшості метаболітів до крові є активним фізіологічним процесом, що потребує затрат енергії макроергічних сполук.
ОСОБЛИВОСТІ ТРАВЛЕННЯ У ПТАХІВ
Основні фізіологічні та біохімічні процеси перетравлювання та усмоктування поживних речовин у птахів подібні до цих процесів у ссавців. Разом з тим, є деякі особливості, що обумовлені різницею у способі життя, характері живлення та морфології травного тракту.
У птахів головними кормами є висококалорійні повнораціонні комбікорми, в основі яких лежать зерноконцентрати з обмеженим набором компонентів рослинного та тваринного походження. До морфологічних
|
особливостей |
травного тракту |
слід |
||||
|
віднести, передусім, відсутність зубів |
||||||
|
та |
доволі |
просту |
|
структуру |
||
|
носоглотки. |
|
|
|
|
||
|
|
Птахи |
захоплюють |
корм |
|||
|
дзьобом, не пережовують його, а |
||||||
|
швидко проковтують. Розм’якшення |
||||||
|
спожитого корму відбувається у волі, |
||||||
|
– випинанні стінки стравоходу. Воло |
||||||
|
добре розвинене у курей, індиків, |
||||||
|
голубів та цесарок. У водоплавних |
||||||
|
птахів справжній зоб відсутній, але у |
||||||
|
кінці |
стравоходу є ампулоподібне |
|||||
|
розширення (несправжній зоб). |
|
|||||
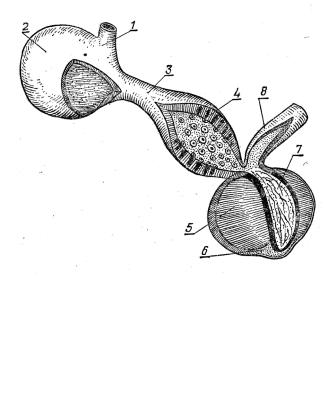
Рис. 22. Будова вола і шлунка птахів: |
|
||||||
|
Шлунок |
складається з |
двох |
||||
1 – стравохід, 2 – воло, 3 – грудна |
відділів – залозистого |
та |
м’язового |
||||
частина стравоходу, 4 – залозистий |
(рис. |
22). |
Обидва |
відділи |
|||
шлунок, 5 – м’язовий шлунок, 6 – |
відокремлені |
один |
від |
одного, |
|||
нижній проміжний м’яз, 7 – кутикула, |
відрізняються |
за |
формою |
та |
|||
8 – дванадцятипала кишка. |
|||||||
функцією. Залозистий відділ шлунка |
|||||||
|
|||||||
за функцією нагадує простий шлунок у ссавців, а м’язовий слугує спеціалізованим органом для перетирання корму.
Залозистий відділ добре розвинений у хижих птахів, а м’язовий – у зерноїдних.
Травлення у волі. У волі є небагато слизових залоз, секрет яких не містить ферментів. Воло – резервуар для корму. Наприклад, у курей у волі розміщується до 100 грамів корму. Тут корм пом’якшується та навіть може частково перетравлюватися під дією ферментів рослинного та бактеріального походження. У волі відбувається гідроліз крохмалю, а також бродильні процеси з утворенням молочної та інших кислот у перші 5 год. після годівлі, а також ліполіз та протеоліз за більш пізнього часу.
Удеяких птахів (голуби) під час вигодовування пташенят у волі утворюється особлива рідина молочного кольору, що містить білки, жири, мінеральні речовини, ферменти та слугує кормом для пташенят.
Загальна тривалість знаходження корму у волі курей, індиків, голубів складає від 3 до 18 год. Під час безперервної годівлі птахів воло звільняється регулярно. Завдяки скороченню мускулатури вола кормова маса із нього прямує до залозистої частини шлунка.
Травлення у шлунку. Шлунок у птахів двокамерний і складається із залозистої та м’язової частини. Слизова оболонка залозистого шлунка вкрита одношаровим циліндричним епітелієм. Тут утворюється шлунковий сік з кислою реакцією, незважаючи на відсутність обкладових клітин.
Усередньому за добу у курей, індиків, гусей та качок виділяється шлунковий сік у кількості 6–9 мл на 1 кг маси за 1 год. Ці покажчики перевищують інтенсивність секреції шлункового соку у людини, собаки та мавпи.
Має місце видова різниця щодо протеолітичної активності та кислотності соку у птахів: найбільшою перетравлюваною силою та кислотністю володіє шлунковий сік курей та індиків, найменшою – гусей.
Сокогінними кормами для птахів є зелені корми, картопля та морква, мучний комбікорм, овес та менш сокогінні – просо, м’ясо-кісткове та рибне борошно.
М’язова частина шлунка складається з двох пар гладеньких м’язів – головних та проміжних. Основна функція м’язового шлунка – стискання та перетирання корму.
Утовщині слизової оболонки м’язового шлунка розміщені довгі та вузькі трубчасті залози, подібні до залоз першого відділу шлунка. Назовні від цього залозистого шару є тверда, зроговіла складчаста оболонка, що має назву кутикула. Кутикула має мукополісахаридну природу та є секретом специфічних
залоз слизової оболонки м язового шлунка.
Ушлунку птахів перетравлюється до 30–50% протеїну, 17–25% вуглеводів та 9–11% жирів, що надходять з кормом.
Травлення у кишках птахів. Травлення в тонких кишках у птахів, так само, як і у ссавців, забезпечується підшлунковим, кишковим соками та жовчю.
Упросвіт 12-палої кишки відкриваються панкреатична та жовчні протоки однією загальною папілою, що стає на заваді одержання чистого підшлункового соку. Секреція панкреатичного соку є безперервною. За добу у
курей виділяється підшлункового соку 27–28 мл на 1 кг маси тіла, у гусей – 15– 20 мл. рН соку – 7,5–8,5, переважно за рахунок гідрокарбонату натрію.
Сік володіє протеолітичною, амілолітичною та ліполітичною активністю. Активність протеаз та амілази значно вищі ніж у ссавців.
Щодо протеїназ, у соку міститься трипсин, хімотрипсин, карбоксипептидази, еластази. У підшлунковому соку також є альфа-амілаза та інвертаза, відсутня лактаза. Секреція соку регулюється такими ж механізмами, що й у ссавців.
Кишковий сік – секрет залозистого епітелію слизової оболонки кишки, виділяється безперервно у кількості 7–9 мл на 1 кг маси тіла курей. У складі кишкового соку є протеїнази, дипептидази, амінопептидази, ентерокіназа, а щодо глюкозидаз, то є амілаза, мальтаза та сахараза, відносно естераз – ліпаза та фосфотази. У кишках птахів основний гідроліз поживних речовин відбувається на поверхні слизової оболонки, тобто за умов контактного травлення. Жовч у птахів як до жовчного міхура, так і безпосередньо до 12палої кишки з правої та лівої долей печінки надходить синусом. Міхурова жовч темно-зеленого кольору, а печінкова – інтенсивно зеленого. На 1 кг маси тіла у птахів жовчі виділяється більше, ніж у інших сільськогосподарських тварин. У голубів та цесарок відсутній жовчний міхур.
Утворення жовчі у гепатоцитах відбувається безперервно, а надходження її до кишки здійснюється рефлекторно. У стані “натщесерце жовч до кишки практично не надходить.
Жовчовиведення регулюється нейрогуморальними механізмами, подібно до ссавців.
Усмоктування в тонких кишках птахів здійснюється за тими ж закономірностями та механізмами, що й у інших видів тварин. Відносно невелика довжина кишкового тракту у птахів та нетривале перебування у ньому корму компенсується інтенсивним перетравлюванням та усмоктуванням продуктів гідролізу.
До товстих кишок птахів належить пряма кишка з парними сліпими відростками. Існують різні погляди щодо ролі сліпих відгалужень. Наявність багаточисельної мікрофлори забезпечує процеси протеолізу, розщеплення клітковини, перетворення небілкових азотистих сполук, синтез вітамінів групи В, що майже усі виділяються з фекаліями.
Контрольні питання:
1.Сутність та призначення травлення.
2.Види та типи травлення.
3.Функції травної системи.
4.Характеристика слинних залоз.
5.Склад та властивості слини.
6.Значення слини.
7.Регуляція слиновиділення.
8.Вплив різних факторів годівлі на кількість та якісний склад слини.