

Физиология Karpovskyi
.pdfЕВАКУАЦІЯ ВМІСТУ ШЛУНКА ДО ТОНКИХ КИШОК
Корм, що надходить до шлунка являє собою суміш рідких та твердих речовин, а вміст шлунка, що залишає шлунок, як правило, має рідку або напіврідку консистенцію. Тривалість перебування корму у шлунку залежить від його складу та консистенції (у середньому 4 10 год.). Вуглеводисті корми переходять до 12-палої кишки швидше, ніж білкові. Повільніше за все зі шлунка евакуюються до кишок жири. Добре подрібнений корм залишає шлунок швидше, ніж погано подрібнений. Ізотонічні розчини переходять до 12-палої кишки швидше, ніж гіпотонічні.
У механізмі евакуації вмісту шлунка до кишок велику роль відіграють: а) тонус пілоричного сфінктера; б) систолічні скорочення антральної частини шлунка, що утворюють градієнт тиску між шлунком та 12-палою кишкою; в) вплив гуморальних факторів регуляції тонусу м’язів шлунка та пілоричного сфінктера; г) функціональний стан центра травлення, що представлений нервовими структурами на різних рівнях центральної нервової системи, включаючи кору великих півкуль головного мозку.
Провідним фактором пілоричного рефлексу є подразнення механота хеморецепторів пілоричної ділянки шлунка (що сприяє евакуації) та рецепторів 12-палої кишки (що гальмує евакуацію). Відомий факт повільного випорожнення вмісту шлунка з високим вмістом жиру обумовлений пригнічуючим впливом ліпідів з 12-палої кишки, а не з самого шлунка. А от постульований Кеноном вплив рН шлункового вмісту на пілоричний сфінктер не знайшов підтвердження у експерименті.
ТРАВЛЕННЯ У ШЛУНКУ КОНЯ
Шлунок коня однокамерний, стравохідно-кишкового типу, невеликий за розмірами. Його об’єм складає 10 15 л. У кардіальній частині шлунка є беззалозиста зона сліпого мішка, устелена багатошаровим плоским епітелієм. Від фундальної та пілоричної зон сліпий мішок відокремлюється вузькою смужкою кардіальних залоз. Стравохід спрямований відносно шлунка під гострим кутом, а функцію кардіального сфінктера виконують дві м’язові петлі, тонус яких підвищується при наповненні шлунка. З цієї причини під час переповнення шлунка у коня стає неможливим акт блювання та відходження газів.
Усі основні механізми секреторної та моторної діяльності шлунка, що встановлені на собаках та інших моногастричних тваринах, притаманні й коню. Разом з тим, є відмітні особливості шлункового травлення у коня:
1. Кінь вельми старанно пережовує корм й тому вміст шлунка має кашкоподібну консистенцію, якою заповнюється, передусім, фундальна ділянка та частина пілоричного відділу шляхом послідовного нашарування вмісту шлунка. У останню чергу заповнюється сліпий мішок. За цих умов було чітко виявлено пошарове розташування порцій корму, вжитого послідовно. Це доведено шляхом огляду шлунка коня після забою тварини.
2.Шлунок коня практично не буває порожнім, незважаючи на те, що його вміст невеликими порціями евакуюється до 12-палої кишки уже невдовзі після початку прийому корму, оскільки через ретельне пережовування вміст шлунка має кашоподібну консистенцію.
3.Завдяки пошаровому розміщенню кормових мас та доволі слабкій моториці шлунка, шлунковий сік повільно просочує шари його вмісту. Зважаючи на це, рН вмісту шлунка у різних шарах та зонах коливається від 1,5
до 4,3, а у ділянці сліпого мішка та у центральній частині шлунка від 6,0 до 6,5. За цих причин у зоні сліпого мішка та у центральній частині шлунка коня мають місце процеси бактеріального бродіння та амілоліз з утворенням молочної кислоти та газів – Н2 та СО2. У невеликих кількостях утворюються оцтова, пропіонова та масляна кислоти. Клітковина у шлунку не перетравлюється через відсутність целюлозолітичної активності.
4.Вода, що споживається конем, не порушує пошарове розміщення спожитого корму та за умов помірного наповнення шлунка швидко прямує до 12-палої кишки в основному ділянкою малої кривизни.
5.Процеси протеолізу спостерігаються, головним чином, у фундальній зоні – у шарах корму, що прилягають до стінки шлунка. Амілолітичні процеси гальмуються при просочуванні шарів корму кислим шлунковим соком. Концентрація НСl у шлунковому соку коня менша ніж у м’ясоїдних та всеїдних
(0,12 0,22%).
6. Секреція шлункового соку є безперервною, незважаючи на те, що інтенсивність її підвищується після годівлі, а також під дією регуляторних факторів. У коней добре виявлена складнорефлекторна, а також шлункова фази секреції. Основним стимулюючим нервом є блукаючий нерв. За умов перерізання обох блукаючих нервів шлункова секреція зменшується до мінімуму, незважаючи на те, що цілком не припиняється через дію гуморальних факторів стимуляції секреції шлункового соку.
ТРАВЛЕННЯ У ШЛУНКУ СВИНІ
Шлунок свиней є однокамерним, шлунково-кишкового типу, ємність його у дорослих тварин складає 0,5 9,0 л. Морфологічна особливість шлунка свині полягає у наявності беззалозистої зони, що охоплює основу стравоходу та малу кривизну шлунка й дивертикулу у кардіальній зоні. Поверхня беззалозистої, кардіальної, фундальної та пілоричної зон складає відповідно 10, 40, 30 та 20% від загальної площі шлунка.
У шлунку свиней кормові маси, так як і у коней, розміщуються пошарово та погано перемішуються. За цих умов у центральній частині шлунка, а також у дивертикулі та верхній частині кардіальної зони відбувається гідроліз крохмалю під дією амілази слини з утворенням мальтози, глюкози та продуктів бактеріального бродіння вуглеводів молочної, оцтової та масляної кислот і газів. У шлунку свиней розщеплюється до 20% легкорозчинних вуглеводів корму. У міру просочування вмісту шлунка кислим шлунковим соком пригнічується процес амілолізу. Величина рН вмісту дивертикула та верхньої
частини кардіальної зони коливається у межах 6,0 7,0, тоді як у шарах, які прилягають до фундальної зони, складає 2,5 3,0.
У шлунковому соку свиней вміст НСl більший, ніж у травоїдних. рН чистого шлункового соку складає 0,7 0,8. У поросят до 30-добового віку у шлунковому соку немає вільної НСl (вікова ахлоргідрія), а кислотність середовища, необхідного для протеолізу забезпечується молочною кислотою.
Під дією пепсинів шлункового соку відбувається перетравлювання білків, а жири гідролізуються дією ліпази, що міститься у кишковому хімусі, який закидається до шлунка.
Секреція шлункового соку у свиней також безперервна, особливо щодо кардіальних залоз. Приймання корму різко підсилює секрецію шлункового соку з добре виявленими фазами секреції складнорефлекторною, шлунковою та кишковою. Кількість шлункового соку залежить від характеру корму, а також частоти годівлі тварин. У середньому за добу у свиней виділяється 4,5 6,0 л. шлункового соку.
Вміст шлунка свиней вже через годину після годівлі починає евакуюватися до 12-палої кишки й за 4 6 год. половина прийнятого корму знаходиться у кишках.
У свиней пілоричний сфінктер, що має вигляд валика, замикається не повністю і тому у процесі антиперистальтики кишок відбувається закидання вмісту кишок (хімусу) до шлунка. Внаслідок цього, за рахунок ліпази кишкового хімусу забезпечуються ліполітичні процеси у шлунку свиней.
Нервово-гуморальна регуляція секреції шлункового соку у свиней відбувається за тими ж механізмами, що й у м’ясоїдних тварин.
ТРАВЛЕННЯ У ШЛУНКУ ЖУЙНИХ ТВАРИН
У жуйних тварин в процесі еволюції сформувався складний, багатокамерний шлунок, що обумовлено, перш за все, характером живлення. Рослинні корми відрізняються низькою поживністю, важко засвоюються та мають невелику біологічну цінність. Тому для забезпечення організму необхідною кількістю та якістю поживних речовин жуйні тварини споживають більшу кількість рослинних кормів, які довгий час знаходяться у травному тракті, де зазнають розпаду та якісних змін.
Шлунок жуйних складається з чотирьох камер рубця, сітки, книжки та сичуга. Перші три камери, що позбавлені залозистої тканини, називають передшлунками, а сичуг є справжнім шлунком через те, що має усі різновидності залозистої тканини подібно шлунку моногастричних. У таких тварин як верблюд, лама, альпака відсутня книжка.
Слизова оболонка передшлунків вистелена багатошаровим плоским епітелієм та має характерну будову у різних камерах: у рубці слизова оболонка має сосочки заввишки від 0,5 до 1 см, причому більш довгими є сосочки у вентральному відділі рубця. У сітці знаходяться комірки, що нагадують бджолині стільники, у книжці листочки різного розміру.
Ємність складного шлунка складає 65 75% загального об’єму травного тракту: від 140 до 300 л. у корови, 15 20 л. у дорослої вівці. Співвідношення ділянок складного шлунка змінюється з віком у міру розвитку передшлунків, причому їх збільшення відбувається нерівномірно. Якщо у новонароджених ємність сичуга у 2 рази більша за об єм передшлунків, то вже у 3-місячному віці передшлунки стають більшими за об єм сичуга майже у 3 рази. Співвідношення передшлунків та сичуга характерним для дорослих тварин є 11:1, що досягається приблизно у 6 7-місячному віці і співпадає з повним становленням травлення у рубці.
Самою місткою камерою є рубець, що займає усю ліву половину черевної порожнини. Інтенсивність розвитку передшлунків з віком залежить, перш за все, від умов живлення. Перехід до кормів, що містять рослинні компоненти у складі стартерів, брикетів, трав’яної муки, гранул, сінажу, сіна та соломи, стимулює розвиток передшлунків. Дослідженням Г.І. Калачнюка та його співробітників було виявлено, що раннє привчання 20-добових бичків до поїдання стартерів спочатку у зволоженому, а потім у сухому вигляді сприяє більш швидкому розвитку передшлунків та становленню рубцевого травлення. Бичкам, що вирощуються за цією технологією, уже у 3-місячному віці вводили до раціону нетрадиційні корми у вигляді сухого пташиного посліду, тирси та небілкових нітрогенвмісних речовин (сечовина, діамонійфосфат). За цих умов стимуляторами росту та розвитку передшлунків і перш за все рубця є не тільки грубі корми, що містять клітковину, але й продукти її мікробіологічної ферментації (оцтова, пропіонова та масляна кислоти). Розвиток сосочків рубця різко збільшує поверхню стінок рубця та площу усмоктування поживних речовин.
Довготривале згодовування жуйним тваринам молока та рідких кормів уповільнює ріст та розвиток передшлунків.
РЕФЛЕКС СТРАВОХІДНОГО ЖОЛОБА
У телят та ягнят під час молочного та перехідного періоду живлення найбільше розвинутою ділянкою складного шлунка є сичуг, у якому починається перетравлення поживних речовин молока та молочних кормів. Надходження прийнятого молока безпосередньо до сичуга забезпечує стравохідний жолоб своєрідне продовження стравоходу.
Стравохідний жолоб являє собою м’язову згортку, за формою у вигляді валиків, що прямує від переддвер’я рубця стінкою сітки до книжки. При цьому, скорочення мускулатури стравохідного жолоба з утворенням своєрідної тимчасової трубки забезпечує надходження рідкого корму, обминаючи рубець та сітку до сичуга. Цей процес є рефлекторним. Рефлекторна дуга рефлексу змикання валиків стравохідного жолоба починається з подразнення рецепторів губ під час смоктання або напування, а також глотки та кореня язика під час ковтання (рис. 10).

Рис. 10. Схема рефлекторної регуляції змикання стравохідного жолоба:
1 – рецептори ротової порожнини; 2 – аферентний шлях від рецепторів ротової порожнини; 3 – центр довгастого мозку; 4 – еферентний шлях, що йде в складі блукаючого нерва; 5 – стравохід; 6 – рубець; 7 – стравохідний жолоб; 8 – сітка; 9 – книжка; 10 – сичуг; 11 – місцезнаходження вищого центру рефлексу стравохідного жолоба у головному мозку
За умови переходу на споживання грубих кормів стінки м’язів стравохідного жолоба грубішають і цей рефлекс проявляється слабше, втрачаючи своє функціональне призначення. У тварин з розвиненими відділами передшлунків як рідкі, так і щільні частинки корму надходять до рубця. Слід відзначити, що за умов вигодовування тварин рідкими кормами, рефлекс стравохідного жолоба зберігається, а розвиток передшлунків загальмовується. Навіть у дорослих тварин можна підтримувати функцію стравохідного жолоба, якщо досить тривало випоювати рідкий корм.
ФУНКЦІЇ ПЕРЕДШЛУНКІВ
Рубець є головним резервуаром корму. Тривалість перебування кормів у рубці складає 3 4 доби і залежить, передусім, від характеру корму. Грубі корми перебувають у рубці значно довше, ніж концентровані та соковиті. Ступінь подрібнення грубих кормів визначає швидкість евакуації вмісту рубця до наступних відділів травного тракту. Так, трав’яна мука, гранульовані корми прямують до сичуга значно раніше, ніж сіно та солома. У рубці корми зазнають фізичних та хімічних перетворень завдяки мікроорганізмам і перемішуються, а при досяганні певної концентрації прямують до сітки. Травлення у рубці частіше всього вивчають за допомогою фістульної методики. Через фістулу, що накладається на рубець у ділянці лівої голодної ямки, можна вивчати моторику балонографічним методом, одержувати проби рубцевого вмісту для аналізів.
Вміст рубця являє собою кашкоподібну масу буро-жовтого або темнозеленого кольору зі своєрідним запахом. Консистенція та склад вмісту рубця залежать від характеру раціону. Вони неоднакові у різних шарах вмісту рубця. Біля основи вентрального мішка, у сітці, а також у верхніх шарах дорсального мішка рубця щільність вмісту менша.
Неоднаковим є у вмісті рубця також співвідношення розмірів кормових частинок, що обумовлено наявністю окремих камер зі своєрідним характером їх слизової оболонки та моторики. Розмір частинок вмісту рубця зменшується від рубця, сітки до сичуга, і звичайно залежить від характеру раціону.
Сітка, маючи коміркову будову слизової оболонки, виконує роль сортувального відділу й пропускає до книжки лише подрібнені частинки корму, а також сторонні предмети, що надходять до рубця під час годівлі тварин. При скороченні сітки, добре подрібнений її вміст прямує до книжки, а більші частинки корму повертаються назад до рубця. Якщо до сітки потрапляють сторонні гострі предмети (дріт, цвяхи), нерідко травмується її стінка, діафрагма та серцева сорочка. За таких умов виникає захворювання травматичний ретикуло-перикардит.
Слизова оболонка книжки являє собою листочки різної висоти, за рахунок цього утворюється гранично велика поверхня для усмоктування. Спочатку вважали, що завдяки листочкам кормові частки перетираються та подрібнюються. Проте електромікроскопічна характеристика гладенької м язової тканини листочків виключає можливості перетирання частин корму. Виявлено, що з порожнини книжки усмоктується до крові велика кількість води, гідрокарбонату натрію, летких жирних кислот. При надходженні до книжки значної кількості концентрованих кормів може виникнути переповнення книжки, бо концентровані корми, що розміщуються поміж листочками, притягують воду, набрякають та стискують листочки.
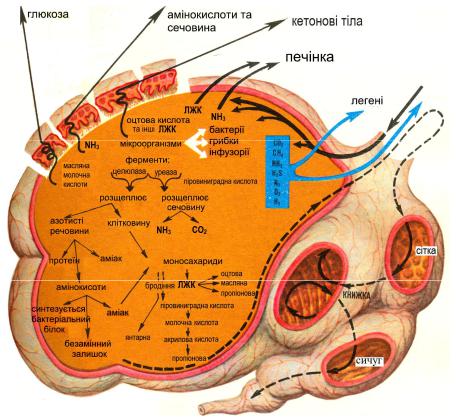
МІКРООРГАНІЗМИ ПЕРЕДШЛУНКІВ ТА ЇХ РОЛЬ У ТРАВЛЕННІ
Відомо, що передшлунки позбавлені залозистої тканини та будь яких травних соків, але в них відбувається перетравлювання всіх поживних речовин: до 70% перетравної клітковини, до 60% перетравного протеїну та до 95% легкорозчинних вуглеводів. Усі найскладніші біохімічні процеси, що проходять у передшлунках, здійснюються ферментами, які виділяються мікроорганізмами, тобто яскраво виявляється так зване симбіонтне травлення. Мікроорганізми рубця – бактерії, інфузорії та гриби. У рубці утворюються сприятливі умови для життєдіяльності мікроорганізмів (стала температура 38–40 С, газовий склад з низьким вмістом О2, оптимальна величина рН, а також постійна наявність субстрату для життєдіяльності мікроорганізмів). У підтриманні відносної сталості вмісту рубця велике значення має двостороння проникність стінки рубця та її метаболічна активність.
Бактерії. У рубці виявлено більше 100 різновидів бактерій. Вже у перші дні життя тварин бактерії надходять до травного тракту з кормом, проте видовий склад їх усталюється вже 1,5 2-місячному віці та залежить, передусім, від складу раціону. Між окремими видами бактерій, як і між бактеріями та
найпростішими, існують особливі форми внутрішніх взаємовідносин (антагонізм, симбіоз та інше). У передшлунках утворюється своя мікробна екологічна система. У цій екосистемі визначають ряд популяцій бактерій: 1) бактерії, що фіксовані на епітелії слизової оболонки рубця (бактерії, що виділяють карбамідамідогідролазу або уреазу та протеолітичні ферменти); 2) бактерії, що фіксовані на поверхні часток корму (целюлозолітичні та амілолітичні бактерії); 3) бактерії, що живуть у рідині рубця (бактерії відповідальні за усі види бродіння).
Ферменти бактерій володіють специфічністю, тобто здатністю діяти на певні поживні речовини. Серед бактерій, що діють на вуглеводи розрізняють: молочнокислі, що зброджують легкорозчинні вуглеводи до молочної кислоти; амілолітичні, що розщепляють крохмаль через стадії декстринів до мальтози, а остання вже гідролізується під дією -глюкозидази до моносахаридів; целюлозолітичні, що гідролізують целюлозу до дисахариду целобіози, що розпадається під дією ферменту целобіази на - та -глюкозу.
До бактерій, що беруть участь у розщепленні білків зараховують: протеолітичні бактерії, що розщеплюють білки до пептидів та амінокислот; декарбоксилюючі бактерії, що відщеплюють СО2 від амінокислот та інших карбонових кислот; дезамінуючі бактерії, що відщеплюють аміногрупу з утворенням амоніаку; бактерії, що виділяють фермент уреазу (карбамідамідогідролазу), під дією якого сечовина гідролізується до NH3 та СО2; бактерії, що здатні синтезувати амінокислоти та білки з використанням нітрогену NH3. Гідроліз ліпідів здійснюється ліполітичними бактеріями з утворенням гліцеролу та карбонових кислот.
Інфузорії в рубці являють собою клас війкових з підкласу рівновійкових (Holotricha) та маловійкових (Oligotrichica) у співвідношенні приблизно 1:9. Визначають більше ніж 50 видів інфузорій рубця. У всіх домашніх жуйних тварин у рубці живе біля 20 видів інфузорій. Загальна маса найпростіших рубця відповідає масі мікрофлори, складаючи приблизно 10% сухої речовини вмісту передшлунків.
Інфузорії рубця беруть участь: у перетравлюванні клітковини механічно подрібнюючи клітковину вони готують її до дії ферментів целюлозолітичних бактерій; у гідролізі крохмалю, а також легкорозчинних вуглеводів. При надмірному надходженні крохмалю з кормом у цитоплазмі інфузорій відкладається до 70% сухої речовини у формі амілопектину. Найпростіші здатні до захоплення бактерій, а володіючи високою протеолітичною активністю вони перетравлюють білки, утворюючи свої специфічні білки, що відрізняються високою біологічною цінністю. Завдяки бактеріям та інфузоріям у рубці синтезується протягом доби до 450 г білка. Біологічна цінність бактерій складає 65%, білка найпростіших – 70%, а їх перетравлювання відповідно 75 та 85%.
Гриби. У рубці виявлена також облігатна мікрофлора анаеробні грибидріжджі, плісняви та деякі види актиноміцетів. Усі вони фіксовані на частках рослинного корму, володіють здатністю гідролізувати клітковину, розщеплювати розчинні вуглеводи, беруть участь у синтезі амінокислот та
вітамінів групи В. Гриби рубця залишаються найменше вивченими у плані участі їх у перетворенні поживних речовин корму.
Слід відзначити, що кількісний та якісний склад мікроорганізмів рубця залежить, перш за все, від характеру раціону, в залежності від складу якого у рубці налагоджується певна мікробна популяція відносно сталого складу. Адаптаційний період для мікроорганізмів рубця за умов зміни раціонів складає у середньому 10 12 діб.
Під час вивчення мікробного складу вмісту рубця слід зважати на те, що мікробна популяція є неоднаковою у різних ділянках рубця та залежить як від характеру живлення, так і часу відбирання проб вмісту рубця після годівлі. Так, у перші 1 2 год. після годівлі у рубці переважають молочнокислі бактерії, оскільки зброджуються легкорозчинні вуглеводи з утворенням молочної кислоти. рН при цьому знижується до 5,5. У ці години є нечисленними інфузорії та малоактивними целюлозолітичні бактерії. Через 6 8 год. після годівлі, коли завершується загалом гідроліз крохмалю, а рН підвищується до 6,5 6,8, у вмісті рубця зростає кількість інфузорій та целюлозолітичних бактерій. Найбільшу кількість інфузорій виявляють у вмісті рубця вранці перед годівлею.
З урахуванням такої динаміки популяцій мікроорганізмів у рубці, як правило, на ніч жуйним тваринам згодовують грубі корми.
ТРАВЛЕННЯ ВУГЛЕВОДІВ У ПЕРЕДШЛУНКАХ
У рослинних кормах вуглеводи складають від 50 до 80% органічних сполук, що являють собою клітковину, геміцелюлозу, інулін, пектинові речовини, крохмаль у складі концентратів, картоплі, а також монота дисахариди. Якщо за будовою усі вуглеводи поділяти на моносахариди (глюкоза, фруктоза, галактоза, маноза), дисахариди (сахароза, лактоза, целобіоза, мальтоза) та полісахариди (целюлоза, крохмаль та інші), то у практиці годівлі тварин доцільніше вуглеводи корму розглядати за ступенем та швидкістю їх засвоюваності.
Легкорозчинні вуглеводи, що надійшли до рубця, зазнають дії молочнокислих бактерій та інфузорій, зброджуються з утворенням молочної кислоти. Вміст останньої у рубці не перевищує 1 2% від загальної кількості кислот бродіння через те, що молочна кислота швидко усмоктується вже у порожнині рубця, а також перетворюється на пропіонову кислоту (рис. 11).
Крохмаль зазнає дії амілолітичних бактерій та інфузорій. Під дією амілази крохмаль гідролізується через стадію декстринів до дисахариду мальтози, а остання розпадається під дією -глюкозидази (мальтази) до двох молекул -глюкози з подальшим бродінням до летких жирних (карбонових) кислот оцтової, пропіонової та масляної. За умов надмірного надходження крохмалю до рубця інфузорії накопичують у цитоплазмі зерна амілопектину, який надалі може бути використаний при дефіциті крохмалю.
Оптимум рН для амілолітичних бактерій складає 5,8 6,2 після того, як молочна кислота (основний продукт ферментації легкорозчинних вуглеводів)
вже всмоктується у кров або перетворюється на пропіонову кислоту. Самим важкозасвоюваним вуглеводом корму є структурний полісахарид рослин
– клітковина або целюлоза.
Геміцелюлоза, лігнін, що містяться у рослинах, до недавнього часу вважалися взагалі
незасвоюваними полісахаридами.
Нещодавно з ґрунтових грибів виділені ферменти, що здатні розщеплювати і ті
компоненти рослинних клітин, що важко перетравлюються. Це дозволило використовувати у раціонах жуйних тварин деревину, стебла бавовни, виноградної лози та інші нетрадиційні корми, що містять клітковину, лігнін та геміцелюлозу.
ЗНАЧЕННЯ КЛІТКОВИНИ. Целюлоза як основний структурний компонент рослин відіграє важливу роль у процесах травлення, виконуючи наступні функції:
стимулятор моторної функції травного тракту як у моногастричних, так і у жуйних тварин. За умов зниження рівня клітковини в раціоні та при сильному подрібненні грубого корму зменшується моторна діяльність рубця, нижчерозміщених ділянок травного тракту через те, що зменшується подразнююча дія грубих часток корму на рецептори травного тракту;
є фактором насичення. Відчуття ситості, насичуваності з’являється за умов певного ступеня наповнення травного резервуару сухою речовиною корму. Клітковина являє собою основну частину сухої частини раціону. Для виникнення такого відчуття ситості у травний тракт лактуючих корів повинно
надходити у середньому 15 17 кг сухої речовини. При згодовуванні тваринам свіжоскошеної зеленої маси з низьким вмістом сухої речовини, тварини споживають надмірну кількість зеленої маси (до 70 кг), що спричиняє нераціональне використання поживних речовин корму. За таких умов посилюється молочнокисле бродіння легкорозчинних вуглеводів, що містяться у зеленій масі, а це в свою чергу спричиняє зниження оцтовокислого бродіння,
а також підвищується рівень пропіонової кислоти. Відомо, що пропіонова кислота є глюкогенною та легко перетворюється у процесі глюконеогенезу на глюкозу й у крові лактуючих корів підвищується концентрація глюкози та знижується концентрація оцтової кислоти. Як наслідок цього, рефлекторно знижується інтенсивність ліполізу у тканинах через підвищений рівень глюкози у крові, а у молоці зменшується вміст жиру через зниження у крові попередників жиру молока та підвищується вміст лактози у молоці;
клітковина виступає також як фактор поверхні. Вище відзначалось, що більша частина бактерій рубця, особливо целюлозолітичних, фіксовані на поверхні грубих частинок корму, тобто клітковини. Через це оптимальний вміст клітковини у раціоні є важливим фактором, що визначає інтенсивність мікробіологічних процесів у передшлунках. Разом з цим необхідно відзначити, що перехід великої кількості клітковини до тонких кишок знижує інтенсивність використання поживних речовин корму, оскільки клітковина має велику поверхню, у кишках адсорбує гідролітичні ферменти, створюючи конкуренцію пристінковому гідролізу поживних речовин;
клітковина як колоїдний субстрат, зв’язує багато води та бере участь у механізмі транспорту її травним трактом;
клітковина належить до числа поганозасвоюваних поживних речовин, складаючи основне джерело сухої речовини рослинних кормів.
Для підвищення ефективності використання клітковини у організмі тварин необхідно піддавати грубі корми відповідному фізичному, хімічному та біологічному обробленню. При фізичному обробленні грубих кормів слід ураховувати видові особливості жуйних тварин. Оптимальними розмірами
подрібнення грубих кормів для корів є 5 7 см, а для овець 2 3 см. При сильному подрібненні грубих кормів, що призначені для лактуючих тварин, спостерігається зниження жирномолочності як результат зниження утворення оцтової кислоти у рубці. Різні види хімічного оброблення грубих кормів, розриваючи хімічні зв язки між перетравленою клітковиною та лігніном рослин сприяють підвищенню перетравності клітковини. Перетравність поживних речовин соломи зростає за умови використання її у процесі силосування кукурудзи, особливо у стадії молочно-воскової стиглості.
Клітковина, що надійшла до рубця, зазнає дії інфузорій, які механічно розпушують клітковину, готуючи ґрунт для целюлозолітичних бактерій. Під дією целюлази клітковина гідролізується повільно до дисахариду целобіози, яка зазнаючи дії ферменту целобіази розпадається до α- та β-глюкози з подальшим бродінням до летких жирних кислот (ЛЖК) – оцтової, пропіонової та масляної з перевагою ацетату.
Таким чином, у жуйних тварин перетворення вуглеводів корму у передшлунках завершується утворенням не глюкози, а летких жирних кислот. У рубці корів утворюється за добу до 5 кг ЛЖК, а у овець – 0,5 кг. Кількість ЛЖК та співвідношення між ними залежать від характеру раціону. Так, за утримання корів на сіно-концентратному раціоні переважає оцтовокисле