

1959 Р – модель Стейна-Даніелі – в біомембранах наявні полярні пори, придатні для трансмембранного переносу гідрофільних молекул.
В 1959 р. Робертсон висунув гіпотезу про унітарний характер біологічних мембран. Зберігаючи трьохшаровий характер для всіх біологічних мембран, він стверджував, що на зовнішній поверхні мембран розташовані також глікопротеїди, тобто мембрана асиметрична за структурою.
В даний час найбільш поширеною є запропонована в 1972 році Синджером і Ніколсоном рідинномозаїчна модель. Ліпіди в мембрані створюють рідку основу, в якій знаходяться рухомі елементи мозаїки - білки.
|
№ п/п |
Назва мембрани |
в % |
||
|
білки |
ліпіди |
вуглеводи |
||
|
1 |
Плазматична мембрана еритроцитів людини |
49 |
43 |
8 |
|
2 |
Внутрішня мембрана мітохондрій печінки |
76 |
24 |
0 |
|
3 |
Мембрана ендоплазматичного ретикулуму клітин печінки |
55 |
45 |
0 |
|
4 |
Мієлінові мембрани мозку людини |
18 |
79 |
3 |
Структурну основу довільної біологічної мембрани складає фосфоліпідний бімолекулярний шар. Він виконує в мембрані функцію бар’єра для іонів і молекул водорозчинних речовин і функцію матриці для мембранних ферментів, рецепторів та інших білків, а також гліколіпідів і глікопротеїдів.
Білки мембран можуть знаходитись на поверхні ліпідного шару, це так звані периферичні білки, або проникати в товщу ліпідного шару, це так звані інтегральні білки. Білки в мембранах утримуються в основному за рахунок електростатичних сил, їх взаємодії з ліпідами. Полярні групи молекул білка напрямлені назовні - в сторону водної фази, а неполярні - в сторону ліпідів. При цьому вважається, що на одну молекулу білка в мембрані припадає 75-90 молекул ліпідів. Оскільки довжина молекули ліпідів дорівнює наближено 3 нм, а товщина моношару білка 1 нм, то товщина клітинної мембрани оцінюється наближено 8 нм.
Білки, які входять до складу мембран, складають 70-75% їх сухої маси. Білки поділяють також на структурні і каталітичні. Останні характеризуються ферментативною активністю.
Другим хімічним компонентом мембран є ліпіди, які складають 20-30% їх сухої маси.
Ліпіди - це група жирів і жироподібних органічних сполук, що мають спільні фізико-хімічні властивості. Ліпіди за хімічним складом поділяють на прості (здебільшого жири) і складні або ліпоїди. Ліпоїди поділяють на фосфоліпіди - сполуки, що містять залишок фосфорної кислоти і гліколіпоїди, що не містять фосфору. Більшість ліпідів мембрани (до 90%) є фосфоліпідами.
Молекулам фосфоліпідів властива амфіфільність, тобто частина молекули (полярна) гідрофільна, а інша – (неполярна) гідрофобна. Це обумовлено особливістю хімічної структури фосфоліпідів, молекула яких містить холін, фосфат і гліцерин, які утворюють полярну “головку”, а також жирні кислоти, які утворюють два неполярні “хвости”. Таку молекулу на схемах позначають:

За формою молекула фосфоліпіду нагадує сплющений циліндр, в якому 1/4 довжини гідрофільна, а 3/4 -гідрофобна.
В водному розчині такі молекули збираються в групи так, що гідрофобні частини молекул закриті від води, а гідрофільні навпаки, контактують з водою. Таким чином, утворюються бімолекулярні шари фосфоліпідів.
ліпосома
Утворення ліпідних мембран, візикул і ліпосом є самодовільним процесом “самозборкою”, обумовленою фізико-хімічною особливістю фосфоліпідних молекул. Бімолекулярні шари фосфоліпідів можуть доповнюватись білками.
Відщеплення одного із жирнокислотних ланцюгів в молекулі фосфоліпіду шляхом окислення або інших процесів приводить до зміни її форми. Така молекула набуває форму конуса, поперечні розміри гідрофобного ланцюга менші від розмірів гідрофільної головки. Такі “дефектні” молекули збираючись разом можуть утворювати пори в бішарі фосфоліпідів.

Діаметр пор у більшості клітин складає 0,35-0,8 нм. Кількість пор в мембрані клітини не велика. В еритроцитах в нормі на пори припадає 0,06% від загальної площі поверхні мембрани. Пори всередині покриті шаром молекул білка. При цьому полярні групи молекул білка направлені в сторону отвору пори, а неполярні взаємодіють з молекулами ліпідів. Полярні групи білків в порах суттєво впливають на проникливість частинок речовини крізь пори.
При великій кількості пор мембрана втрачає свої бар’єрні властивості. До появи нових пор приводить окислення ліпідів і дія фосфоліпаз, дані процеси мають місце при ряді захворювань.
Пори в біології і фізіології часто називають каналами.
Ліпіди в мембранах мають велику рухливість, яка проявляється в хаотичному тепловому русі молекул ліпідів і білків в площині мембрани. Такий рух називається латеральною дифузією (вздовж біліпідного шару). Швидкість латеральної дифузії досить значна. (Фліп-флоп – дифузія впоперек). Молекули фосфоліпідів здійснюючи до 107 перескоків (обмінів місцями) за секунду і можуть проходити відстань біля 5 мкм. Білки можуть проходити відстань 0,2 мкм за 1 с. Молекули білків і фосфоліпідів зазнають також обертальних рухів. За час 10-9 - 10-4 с вони здатні повертатись на кут до 1-го радіана (570).
Таким чином, при фізіологічних температурах мембрана нагадує бушуюче море, на поверхні якого переміщуються, обертаються, зазнають зіткнень білкові “судна”.
Латеральна дифузія забезпечує велику швидкість ферментативних та інших хімічних процесів, які протікають на поверхні мембрани.
Перехід молекул фосфоліпідів (а тим більше білків) з одної поверхні мембрани на іншу трансмембранний перехід малоймовірний. Перехід молекули фосфоліпіду на протилежну поверхню мембрани відбувається в середньому лише один раз протягом декількох годин. Завдяки цьому мембрани асиметричні, тобто внутрішня і зовнішня поверхні всіх мембран мають різний ліпідний і білковий склад. Асиметрія відіграє важливу роль в транспорті речовин через мембрану.
Модельні ліпідні мембрани
При вивченні будови і функціонування біологічних мембран, механізмів порушення бар’єрних і структурних властивостей ліпідного шару, дії лікарських препаратів, важливу роль відіграють модельні мембрани або штучні мембрани. Техніка створення штучних ліпідних мембран була розроблена Лангмюром в 1917 р.
Найпростішою моделлю є моношар фосфоліпідів на межі води і повітря або води і олії.

На таких межах молекули фосфоліпідів розташовані так, що гідрофільні головки знаходяться у воді, а гідрофобні “хвости” у повітрі або олії. Якщо поступово зменшувати площу, яку займає моношар, то в кінці кінців вдається отримати моношар, в якому молекули розташовані так само щільно, як і в одному із бішарів мембрани.
Мономолекулярний шар фосфогліцериду дипольмітоїлфосфатидилхомін вистилає легеневі альвеоли, протидіє їх злипанню.
Площа поверхні, яка припадає на одну молекулу фосфоліпіда в залежності від стану жирних кислот, складає 0,6 - 0,8 нм2.
Зміни стану молекул ліпідів пов’язані із зміною температури, хімічною модифікацією жирнокислотних “хвостів”, із змінами електричного заряду головки, супроводжуються змінами площі, яка припадає на одну молекулу. Аналогічне явище спостерігається при взаємодії мембран з лікарськими препаратами, наприклад, з анестетиками. Тому вивчення моношарів фосфоліпідів і ліпідів виділених із мембранних структур нормальної і патологічно зміненої клітини є важливим в біологічних і медичних дослідженнях.
Другою моделлю є плоскі біліпідні мембрани. Вперше така модельна мембрана була створена в 1962 р. П.Мюлером і співробітниками. Вони заповнили маленький отвір (діаметром біля 1 мм) в тефлоновій перегородці, яка розділяла два водних розчини, фосфоліпідами, розчиненими у гептані. Після того, як розчинник і надлишки ліпідів розтікались по тефлону, в отворі утворювався бішар товщиною 5-7 нм. Розташувавши по два боки такої мембрани мікроелектроди можна виміряти електричний опір мембрани, або електричну різницю потенціалів, яка виникає на мембрані, коли навколо неї знаходятся розчини різних концентрацій. Такі моделі дозволили вивчити іонну проникливість мембран.
Третьою модельною системою є ліпосоми. Так називають фосфоліпідні бульбашки, які можна отримати під дією ультразвуку, або механічним струшуванням сухих фосфоліпідів у водно-сольовому буфері. При цьому можуть утворюватись одно- і двошарові ліпосоми. Ліпосоми нагадують біологічні мембрани, які позбавлені білкових молекул. Якщо ліпосоми приготувати в середовищі, яке містить досліджувану речовину, а потім змінити концентрацію даної речовини в оточуючому ліпосоми середовищі, то можна кількісно вивчити проникливість ліпідного шару для даної сполуки.
Фізичний стан ліпідів
Вивчення стану фосфоліпідів в модельних і біологічних мембранах проводиться за допомогою методів: рентгеноструктурного аналізу, радіоспектроскопії, дослідження інфрачервоних спектрів поглинання, спектрів комбінаційного розсіювання, люмінесценції, мікрокалориметрії, методів ЕПР та ЯМР та інших.
Дані, отримані цими методами, вказують на те, що бішар ліпідів в біологічних і модельних мембранах може знаходитись в двох станах: твердокристалічному або рідіннокристалічному.
В твердому і рідкому станах зберігається бімолекулярна структура мембрани і щільна гексогональна упаковка молекул фосфоліпідів в площині мембрани. Площа, яка припадає на одну молекулу фосфоліпіду в залежності від структури жирних кислот в твердому стані 0,46-0,48 нм2, а в рідкому 0,6-0,8 нм2. Товщина бішару в рідкому стані менша ніж в твердому.
Рідкий і твердий стани бішару відрізняються в’язкістю ліпідної фази і розчинністю різних речовин в ліпідній фазі.
Молекулярна основа таких відмінностей полягає в конформації жирнокислотних ланцюгів. Окремий жирнокислотний ланцюг може приймати різні конформації завдяки можливості обертання навколо одинарних С-С зв’язків. В щільній упаковці молекул в ліпідному бішарі здебільшого реалізуються дві конформації вуглеводневих ланцюгів жирних кислот:
повна трансконформація
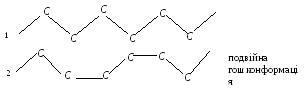
В твердому стані молекули фосфоліпідів мають повну трансконформацію вуглеводневих ланцюгів жирних кислот. Рухливість таких ланцюгів обмежена: вони можуть здійснювати узгоджені коливальні рухи і обертальні рухи навколо точки прикріплення жирних кислот до полярної головки.
В рідкому стані можливі теплові рухи жирнокислотних ланцюгів, які супроводжуються транс-гошпереходами.
Очевидно, що, чим вища рухливість жирнокислотних ланцюгів в мембрані, тим швидше дифундують молекули речовин через ліпідний бішар, тобто тим менша мікров’язкість мембрани.
Співвідношення між рідкою і твердою фазами в ліпідному бішарі залежить від хімічного складу ліпідів, числа заряджених груп, які припадають на одиницю площі поверхні мембрани і від температури.
Температура, при якій половина молекул фосфоліпідів мембрани знаходиться в твердому стані, а друга половина в рідкому, називається температурою фазового переходу, або температурою плавлення.
Для біологічних мембран температура фазового переходу, знаходиться в області від’ємних температур за шкалою Цельсія. Різні за хімічним складом фосфоліпіди мають різну температуру фазового переходу.
При поступовому охолодженні органів, тканин і окремих клітин з метою їх консервації до температури -10˚С, -30˚С в мембранах відбувається послідовне “виморожування” тих ліпідів, температура фазового переходу яких вища температури оточуючого середовища. При цьому, розчинені в ліпідній фазі низькомолекулярні сполуки і білки витісняються в рідіннокристалічні ділянки мембрани. Тому при повній кристалізації ліпідного шару можуть відбутись незворотні зміни в структурі мембрани. Це є один із факторів, які знижують життєздатність клітин при консервації клітин і тканин методом глибокого охолодження.
Явища переносу. Дифузія. Рівняння Фіка
Важливим елементом у функціонуванні біологічних мембран є перенос або транспорт речовин.
Транспорт речовин крізь мембрани поділяють на пасивний і активний.
Пасивним транспортом називають перенос речовини через мембрану за рахунок енергії градієнтів: 1) концентрації речовини (дифузія), 2) електричного потенціалу (електродифузія), 3) осмотичного тиску (осмос), 4) гідростатичного тиску (фільтрація).
Активним транспортом називають перенос речовини через мембрану за рахунок енергії макроергічних зв’язків АТФ. Він здійснюється в напрямі протилежному до дифузії.
Дифузія – лат. diffusio – поширення – взаємне проникнення речовин при безпосередньому стиканні або крізь порувату перегородку, зумовлене тепловим рухом атомів чи молекул речовин.
При звичайній дифузії, тобто переносі незаряджених частинок через мембрану, маса речовини, яка переноситься через перпендикулярну до потоку поверхню мембрани площою S, за час dt, визначається за формулою:
![]()
де D - коефіцієнт дифузії, який залежить від природи речовини і температури;
![]() градієнт
концентрації речовини.
градієнт
концентрації речовини.
Знак “ – “ вказує на те, що речовина переноситься в область меншої концентрації речовини.