

3.3. Гістогенез крові
Кровотворна система в цілому це — відкрита система, що знаходиться у стаціонарному стані, при якому в ній підтримується певне співвідношення між різними клітинними популяціями.
Кровотворення у дорослому організмі вищих хребетних відбувається в різних органах. В епіфізах трубчастих кісток, кістках грудини і черепа здійснюється утворення еритроцитів, зернистих лейкоцитів, кров'яних пластинок, моноцитів і частини лімфоцитів. Крім того утворення лімфоцитів відбувається у селезінці, лімфовузлах, тимусі, мигдалинах і апендиксі.
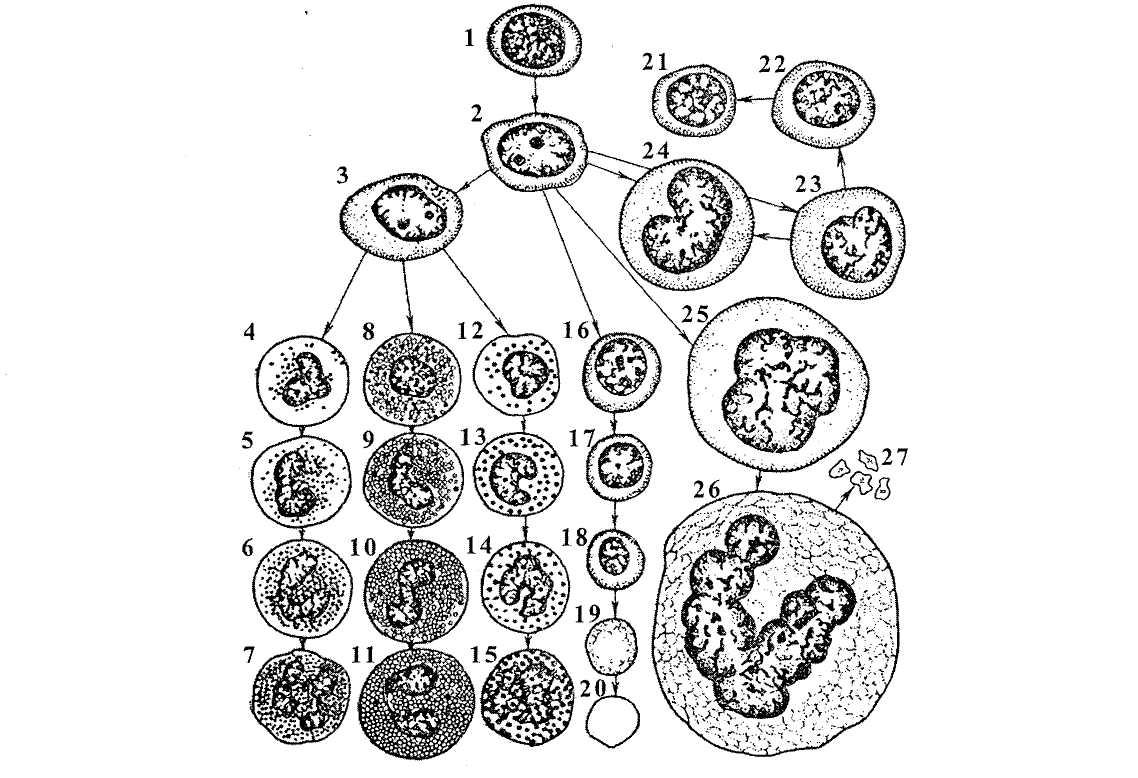
Рис. 24. Етапи кровотворення в дорослому організмі: 1 - стовбурова гіпотетична клітина; 2 - гемоцитобласт; 3 - промієлоцит; 4 - нейтрофільний
46
мієлоцит; 5 - юний нейтрофіл (метамієлоцит); 6 - паличкоядерний нейтрофіл; 7 - сегментоядерний нейтрофіл; 8 - еозинофільний мієлоцит; 9 - юний еозинофіл (метамієлоцит); 10 - паличкоядерний еозинофіл; 11 - зрілий (сегментоядерний) еозинофіл; 12 - базофільний мієлоцит; 13 - юний базофіл; 14 - паличкоядерний базофіл; 15 - зрілий (сегментоядерний) базофіл; 16 - базофільний еритробласт; 17 - поліхроматофільний еритробласт; 18 - нормобласт; 19 - ретиколоцит; 20 -еритроцит; 21-23 -малий, середній і великий лімфоцит; 24 - моноцит; 25 -промегакаріоцит (мегакаріобласт); 26 - мегакаріоцит; 27 - кров'яні пластинки.
Зазвичай у підручниках гістогенез крові викладений за схемою, запропонованою О. О. Максимовим, котрий вважав вихідною клітиною для всіх гілок кровотворення гемоцитобласт — велику базофільну клітину з круглим, або овальним світлим ядром і кількома великими ядерцями. Жодних специфічних гранул в її цитоплазмі немає.
Поділяючись і диференціюючись, ця клітина проходить через ряд проміжних стадій ' і перетворюється в еритроцити, різні гранулоцити, агранулоцити і кров'яні пластинки. Свою унітарну теорію кровотворення О. О. Максимов сформулював 90 років тому. У принциповому відношенні ця теорія в наш час є найбільш імовірною, хоча в окремих ланках не повністю вивчена.
Найбільш суттєвим відкриттям в проблемі кровотворення після робіт Максимова можна вважати: 1 - встановлення факту, що вихідною кров'яною клітиною є не гемоцитобласт, а ще менш диференційована "стовбурова" клітина; 2 - відкриття факторів, що індукують диференціювання стовбурових клітин в тому, чи іншому напрямку (еритропоетин, гранулопоетичний індуктор, тромбоцитопоетин); 3 - показана роль тимусу і антигенів у диференціюванні і проліферації елементів лімфоїдного ряду.
3.3.1. Еритропоез
Розвиток еритроцитів (еритропоез) у дорослому організмі ссавців відбувається в основному в червоному кістковому мозку (у гризунів також в 47
селезінці).
Еритропоез починається зі стовбурової клітини. Потомки цієї клітини — гемоцитобласти. В ході гістогенезу гемоцитобласт стає чутливим до дії гормону еритропоетину. Деякі автори таку гемопоетинчутливу групу гемоцитобластів виділяють у самостійний клітинний тип. Ці клітини здатні до поділу і диференціюються в еритробласти. Ці клітини мають щільне ядро і базофільну інтенсивно забарвлену цитоплазму. Навколо ядра спостерігається вузький обідок світлої перинуклеарної цитоплазми. Вони поступово нагромаджують гемоглобін. Про еритробласти діляться і диференціюються у базофільні еритробласти, для яких характерна висока базофілія цитоплазми, наявність ексцентрично розміщеного колесоподібного ядра. Розміри цих клітин дещо менші, ніж в проеритробластів.
В міру нагромадження гемоглобіну базофілія цитоплазми у базофільних еритробластах починає слабша і вони перетворюються у поліхроматофільні еритробласти. Це клітини, що мають типові колесоподібні ядра, а цитоплазма, завдяки нагромадженню в ній оксифільного гемоглобіну, починає фарбуватися і основним і кислим барвниками одночасно (поліхромно).
Поліхроматофільні еритробласти діляться і диференціюються на оксифільні еритробласти (часто їх називають нормобластами). Ці клітини мають оксифільну цитоплазму, багато гемоглобіну, мало рибосом, мітохондрій та інших органоїдів. Клітинний центр у них зникає і нормобласти стають нездатними до поділу. Ядра цих клітин дрібні і сильно пікнотизовані.
Нормобласти в ретикулоцити. Цей перехід здійснюється шляхом відривання ядровмісного фрагменту від оксифільного еритробласта. На електронних мікрофотографіях видно, що ядерний фрагмент нормобласта оточений тонким шаром цитоплазми. Ці спостереження дозволяють відмовитися від старого уявлення про перехід нормобласта в ретикулоцит, як процес виштовхування ядра з клітини.
У ретикулоциті деякий час зберігаються залишки РНК у вигляді тонкої сіточки - "ретикулюма" (звідси і назва клітини), невелика кількість полісом та ендоплазматичної сітки. Поступово ці клітинні структури зникають, процес 48
синтезу гемоглобіну зупиняється і ретикулоцит стає еритроцитом.
Вивчення кінетики еритропоезу показало, що за нормальних умов для підтримання стаціонарного стану еритроїдної системи відбувається фізіологічна регенерація за рахунок поділу юних (базофільних та поліхроматофільних) еритробластів. В експериментальних умовах (кровотеча, гемоліз) у процес диференціації включаються проеритробласти і гемоцитобласти.