

Обратимые ингибиторы могут быть конкурентными и неконкурентными
Различия между этими типами обратимых ингибиторов касаются их взаимоотношений с субстратом. Если ингибитор конкурирует с субстратом за место связывания на ферменте и действие одного может быть отменено избытком другого, говорят о конкурентном торможении, в противном случае речь идет о неконкурентных ингибиторах.
Конкурентные ингибиторы не всегда структурно подобны субстрату.
Различают два механизма конкурентного торможения.
А.Конкурентное торможение путем связывания активного центра. Классический конкурентный ингибитор - вещество, которое имеет структурное сходство с субстратом фермента. Благодаря этому подобию ингибитор может связываться с активным центром вместо субстрата. Это своеобразная молекулярная ошибка. Однако, поскольку субстрат и ингибитор не идентичны полностью, фермент не способен катализировать превращение ингибитора в продукт. Ингибитор просто блокирует активный центр фермента. Если субстрат свяжется с активным центром раньше, чем ингибитор, ингибитор не может связаться с ферментом. Нельзя одновременно обоим связаться с активным центром. Такой способ конкурентного торможения получил название изостерического из-за схожести (изос) структур субстрата и ингибитора. Наиболее часто в клетке в роли классического конкурентного ингибитора выступает продукт данной реакции, что имеет глубокий практический смысл.
 Б.
Конкурентное торможение путем изменения
конформации фермента. В
отличие от классического варианта,
ингибитор связывается не с активным
центром, а со специальным центром,
связывающим ингибитор, который расположен
вдали от активного центра. Связывание
ингибитора вызывает изменение
пространственной структуры (изменение
конформации) в области активного центра,
которое не позволяет присоединиться
субстрату. Предшествующее связывание
субстрата к активному центру в свою
очередь, вызывает изменения конформации
центра связывания ингибитора, которое
предотвращает связывание ингибитора.
И субстрат и ингибитор не могут
одновременно связаться с ферментом. В
этом виде конкурентного торможения
ингибитор может иметь любую химическую
структуру, поскольку они связываются
с различными участками фермента.
Б.
Конкурентное торможение путем изменения
конформации фермента. В
отличие от классического варианта,
ингибитор связывается не с активным
центром, а со специальным центром,
связывающим ингибитор, который расположен
вдали от активного центра. Связывание
ингибитора вызывает изменение
пространственной структуры (изменение
конформации) в области активного центра,
которое не позволяет присоединиться
субстрату. Предшествующее связывание
субстрата к активному центру в свою
очередь, вызывает изменения конформации
центра связывания ингибитора, которое
предотвращает связывание ингибитора.
И субстрат и ингибитор не могут
одновременно связаться с ферментом. В
этом виде конкурентного торможения
ингибитор может иметь любую химическую
структуру, поскольку они связываются
с различными участками фермента.
Конкурентные ингибиторы не влияют на Vmax, они понижают Км.
Реакции связывания и субстрата и
конкурентного ингибитора протекают
быстро и обратимы, так что они существуют
в равновесии. Позиции этого равновесия
будут зависеть от концентраций реагентов.
Учитывая, что фермент участник обеих
реакций их равновесн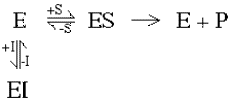 ые
состояния взаимосвязаны. Это означает,
что при высоких уровнях ингибитора
фактически все молекулы фермента будут
участвовать в образованииEIкомплекса, и фермент будет почти полностью
ингибирован. С другой стороны, при
высоких концентрациях субстрата почти
все молекулы фермента будут связаны
вESкомплексе и ингибитор
не сможет связаться с ферментом. Высокие
концентрации субстрата снимают действие
ингибитора. Субстрат и ингибитор
конкурируют друг с другом. Какое влияние
это будет иметь на кинетике (рис 2-10)?
ые
состояния взаимосвязаны. Это означает,
что при высоких уровнях ингибитора
фактически все молекулы фермента будут
участвовать в образованииEIкомплекса, и фермент будет почти полностью
ингибирован. С другой стороны, при
высоких концентрациях субстрата почти
все молекулы фермента будут связаны
вESкомплексе и ингибитор
не сможет связаться с ферментом. Высокие
концентрации субстрата снимают действие
ингибитора. Субстрат и ингибитор
конкурируют друг с другом. Какое влияние
это будет иметь на кинетике (рис 2-10)?

Рис 2-10. График Лаинуивера-Берка в случае классического конкурентного ингибирования
Эффект на KmКм - индикатор сродства субстрата и фермента. В присутствии конкурентного ингибитора некоторые молекулы фермента будут существовать как свободные ферменты, другие как комплексы ингибитора фермента. Первые будут иметь нормальное сродство, а вторые полностью неспособны к связыванию субстрата. Км измеряет полное сродство фермента в реагирующей смеси, которое будет представлять среднее значение между нормой и нулевым значением этого сродства, и поэтому будет явно меньше нормального значения. Так что конкурентный ингибитор уменьшает сродство субстрата и фермента, или увеличивает Км.
Эффект на Vmax Vmax - скорость при высоких концентрациях субстрата. Поскольку в этих условиях, ингибитор вытесняется субстратом, он не тормозит фермент вообще и, следовательно, конкурентные ингибиторы не замедляют реакцию при высоких концентрациях субстрата, и не изменяют Vmax. Это можно хорошо видеть на графике Лайнуивера-Берка. Наклон графика равен Km/Vmax. Увеличение наклона в присутствии ингибитора указывает на снижение скорости реакции при низких уровнях субстрата.
Наиболее часто приводимый пример конкурентного ингибирования - это использование малоновой кислоты для торможение дегидрогеназы янтарной кислоты. Как видно из рисунка 2-9 наиболее близким структурным аналогом сукцината является малоновая кислота.
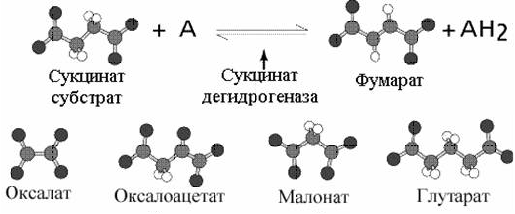
Рис 2-9. Примеры конкурентных ингибиторов.
Малоновая кислота тормозит активность дегидрогеназы янтарной кислоты, занимая активный центр на ферменте. Учитывая обратимость реакции, избыток янтарной кислоты снимет действие малоновой кислоты.