

Министерство здравоохранения Республики Беларусь
Учреждение образования
«Гомельский государственный медицинский университет»
Кафедра биологической химии
Обсуждено на заседании кафедры биологической химии
Протокол № __________
Методическая разработка
Для проведения занятия со студентами
2 Курса фпсзс по биологической химии
(наименование дисциплины)
Тема: Белки 3. Особенности обмена аминокислот в норме и при патологии.
Время 3ч.
1. УЧЕБНЫЕ И ВОСПИТАТЕЛЬНЫЕ ЦЕЛИ, МОТИВАЦИЯ ДЛЯ УСВОЕНИЯ ТЕМЫ; ТРЕБОВАНИЕ К ИСХОДНОМУ УРОВНЮ ЗНАНИЙ.
Цель занятия: сформировать представления об особенностях обмена отдельных аминокислот (АК) в норме и при патологии. Дать биохимическое обоснование практического применения аминокислот в медицине. Освоить методику определения активности трансаминаз в сыворотке крови.
В результате проведения занятия студент должен:
1) Знать специфические пути метаболизма протеиногенных аминокислот, их биомедицинское значение.
2) Научиться проводить исследование на колориметре.
2. КОНТРОЛЬНЫЕ ВОПРОСЫ ИЗ СМЕЖНЫХ ДИСЦИПЛИН.
2.1. Строение, классификация и химические свойства аминокислот (биоорганическая химия).
3.КОНТРОЛЬНЫЕ ВОПРОСЫ ПО ТЕМЕ ЗАНЯТИЯ.
3.1. Цтк (реакции, ферменты, коферменты, механизмы регуляции, биологическая роль). Пути вступления отдельных аминокислот в цтк (глико- и кетогенные аминокислоты).
Цикл трикарбоновых кислот (ЦТК) впервые был открыт английским биохимиком Г. Кребсом. Он первым постулировал значение данного цикла для полного сгорания пирувата, главным источником которого является гликолитическое превращение углеводов. ЦТК является тем центром, в котором сходятся практически все метаболические пути. Таким образом, цикл Кребса – общий конечный путь окисления ацетильных групп (в виде ацетил-КоА), в которые превращается в процессе катаболизма большая часть органических молекул, играющих роль «клеточного топлива»: углеводов, жирных кислот и аминокислот.
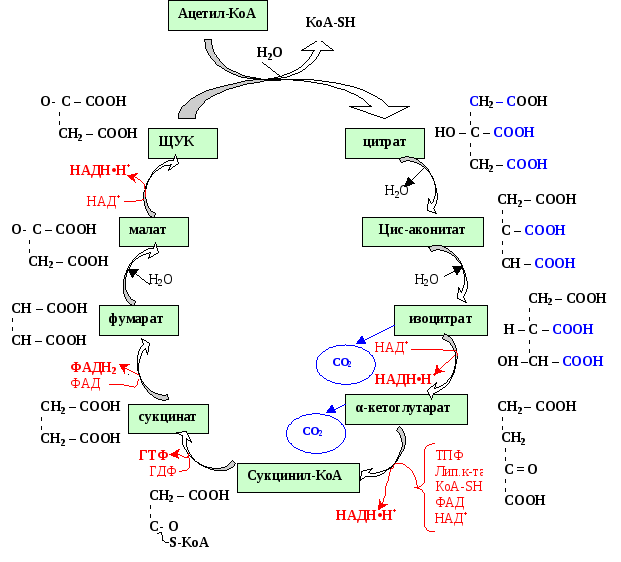
ЦТК происходит в матриксе митохондрий и состоит из восьми последовательных реакций. Начинается цикл с присоединения ацетил-КоА к оксалоацетату и образования лимонной кислоты (цитрата). Затем лимонная кислота (шестиуглеродное соединение) путем ряда дегидрирований (отнятие водорода) и двух декарбоксилирований (отщепление СО2) теряет два углеродных атома и снова в цикле Кребса превращается в оксалоацетат (четырехуглеродное соединение), т.е. в результате полного оборота цикла одна молекула ацетил-КоА сгорает до СО2 и Н2О, а молекула оксалоацетата регенерируется.
Механизмы регуляции:
Скорость реакция цикла Кребса определяется энергетическими потребностями клетки.
Непосредственно регулируется активность дегидрогеназ цикла Кребса, которая зависит от величины отношения НАДН∙Н+/ НАД+: при недостатке НАД+ активность дегидрогеназ снижается, поскольку именно окисленная форма кофермента является акцептором водородов для субстратов ЦТК.
Помимо этого, существует аллостерический механизм регуляции: для изоцитрадегидрогеназы аллостерическими ингибиторами являются АТФ и НАДН∙Н+, активатором – АДФ. Сукцинатдегидрогеназа угнетается избытком субстрата.
Биологическая роль.
Интегративная – ЦТК является центральным метаболическим путём, объединяющим процессы распада и синтеза важнейших компонентов клетки.
Анаболическая – субстраты цикла используются для синтеза многих других соединений: оксалацетат используется для синтеза глюкозы (глюконеогенез) и синтеза аспарагиновой кислоты, ацетил-КоА – для синтеза гема, α-кетоглутарат – для синтеза глутаминовой кислоты, ацетил-КоА – для синтеза жирных кислот, холестерола, стероидных гормонов, ацетоновых тел и др.
Катаболическая – в этом цикле завершают свой путь продукты распада глюкозы, жирных кислот, кетогенных аминокислот – все они превращаются в ацетил-КоА; глутаминовая кислота – в α-кетоглутаровую; аспарагиновая – в ЩУК и пр.
Собственно энергетическая – одна из реакций цикла (распад сукцинил-КоА) является реакцией субстратного фосфорилирования. В ходе этой реакции образуется 1 молекула ГТФ (реакция перефосфорилирования приводит к образованию АТФ).
Водороддонорная – при окислении 3-х НАД+-зависимых субстратов (изоцитрата, α-кетоглутарата и малата) и ФАД-зависимого сукцината образуются 3 НАДН∙Н+ и 1 ФАДН2. Эти восстановленные коферменты являются донорами водорода для дыхательной цепи митохондрий, энергия переноса водородов используется для синтеза АТФ. При этом за счёт окисления 3-х молекул НАДН∙Н+ образуется 3АТФ × 3 = 9 АТФ и за счёт окисления 1 молекулы ФАДН2 - 2 АТФ. Итого водороддонорная функция цикла Кребса обеспечивает образование 11 молекул АТФ, а, учитывая энергетическую функцию цикла, получаем за полный его оборот («сгорание» 1 молекулы ацетил-КоА) 12 молекул АТФ.
Анаплеротическая – восполняющая. Основным дефицитом при работе цикла является ЩУК. Её недостаток восполняется реакцией, катализируемой пируваткарбоксилазой:
Пируват-
карбоксилаза
П ВК
+ НСО3-
– биотин + Н2О
ЩУК + АДФ + Н3РО4
ВК
+ НСО3-
– биотин + Н2О
ЩУК + АДФ + Н3РО4
Пути вступления отдельных аминокислот в ЦТК (глико- и кетогенные аминокислоты).
См тему «Белки-2» вопрос № 1.4