

Кинетика мультисубстратных реакций
Большинство биохимических реакций в клетке протекает с участием двух или более молекул субстрата и с образованием нескольких продуктов. Например, в реакции гидролиза белка участвуют два субстрата (полипептид и вода) и образуются два продукта (два фрагмента расщепленной полипептидной цепочки). Кинетика мультисубстратных реакции несколько сложнее чем кинетика фермента с одним субстратом. Однако, для многих целей, в частности для лабораторного анализа фермента, достаточно определить Км для каждого субстрата при насыщенных концентрациях этих субстратов. Кроме того, принципы кинетики простых реакций могут быть распространены и на полисубстратные реакции, например такие как реакции переноса групп. Это делается путем измерения эффектов различных концентрации одного субстрата на кинетические результаты, полученные для другого.
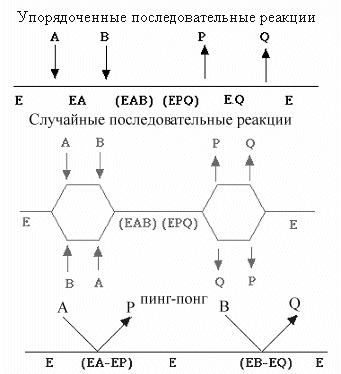 Полисубстратные
реакции можно описывать используя
несколько различных кинетических схем,
называемых кинетическими механизмами.
Кинетические механизмы обычно
представляются, используя систему
обозначений, введенную Клелэндом
(Cleland
W.W.).
Последовательность шагов изображают
слева направо. Добавление молекул
субстрата (A,
B,
C)
к ферменту и отделение продуктов (P,
Q,
R)
от фермента обозначено стрелками.
Различные формы фермента (свободный E,
ES
комплексы, или EP
комплексы) написаны под горизонтальной
строкой. ES
комплексы, которые подвергаются
химическому преобразованию, занимая
активный центр, записываются в круглых
скобках. Последовательные реакции - это
реакции которые требуют, чтобы все
субстраты присутствовали перед тем как
будет образован любой продукт.
Последовательные реакции могут быть
или упорядоченными, при которых
соблюдается обязательный порядок
добавления субстратов и высвобождения
продуктов, или случайными, при которых
не соблюдается обязательный порядок
связывания или высвобождения. В реакциях
пинг-понга продукт высвобождается
раньше, чем свяжутся все субстраты. Для
бисубстратных реакций этого типа,
после связывания первого субстрата
фермент модифицирует свою структуру,
и катализирует образование первого
продукта, после чего связывается второй
субстрат и измененный фермент
восстанавливается в свою первоначальную
форму и высвобождает второй продукт.
Связывание и освобождение лигандов в
пинг-понговом механизме обычно указывается
наклонными линиями.
Полисубстратные
реакции можно описывать используя
несколько различных кинетических схем,
называемых кинетическими механизмами.
Кинетические механизмы обычно
представляются, используя систему
обозначений, введенную Клелэндом
(Cleland
W.W.).
Последовательность шагов изображают
слева направо. Добавление молекул
субстрата (A,
B,
C)
к ферменту и отделение продуктов (P,
Q,
R)
от фермента обозначено стрелками.
Различные формы фермента (свободный E,
ES
комплексы, или EP
комплексы) написаны под горизонтальной
строкой. ES
комплексы, которые подвергаются
химическому преобразованию, занимая
активный центр, записываются в круглых
скобках. Последовательные реакции - это
реакции которые требуют, чтобы все
субстраты присутствовали перед тем как
будет образован любой продукт.
Последовательные реакции могут быть
или упорядоченными, при которых
соблюдается обязательный порядок
добавления субстратов и высвобождения
продуктов, или случайными, при которых
не соблюдается обязательный порядок
связывания или высвобождения. В реакциях
пинг-понга продукт высвобождается
раньше, чем свяжутся все субстраты. Для
бисубстратных реакций этого типа,
после связывания первого субстрата
фермент модифицирует свою структуру,
и катализирует образование первого
продукта, после чего связывается второй
субстрат и измененный фермент
восстанавливается в свою первоначальную
форму и высвобождает второй продукт.
Связывание и освобождение лигандов в
пинг-понговом механизме обычно указывается
наклонными линиями.
Скорость реакции зависит от концентрации фермента
Концентрация фермента также влияет на скорость химической реакции. На рис.2-9 показано влияние снижения концентрации фермента наполовину от исходной на значения двух кинетических параметров Vmax и Км при насыщающей концентрации субстрата. В этих условиях, каждая молекула фермента связана с молекулой субстрата и будет взаимодействовать с ним, превращая его в продукт, со скоростью какой только может. Если удвоить число молекул фермента, мы получим в два раза больше ферментов, связанных с субстратом, так что скорость суммарной реакции должна удвоиться

Рис 2-9.Зависимость скорости реакции от концентрации фермента. Используемые концентрации фермента различались в два раза (см текст).
Из графика видно, что Vmax реакции снизилась с 10 до 5 единиц. Значение Км при этом не изменилось. Изменение числа молекул фермента, таким образом, не изменило их индивидуальных химических характеристик, и сродство фермента к субстрату осталось прежним, хотя максимальная скорость при этом пропорциональна количеству фермента. Это обстоятельство находит широкое применение для исследования количества ферментов в клинико-биохимической диагностике патологических состояний, при которых ферменты из повреждённых тканей попадают в кровь.
Исследование обычно проводится в условиях максимального насыщения ферментов субстратами, поскольку в таких условиях значительные колебания концентрации субстрата не оказывают влияние на измеряемую активность. Это зона реакции нулевого порядка на графике зависимости скорости реакции от количества субстрата. Однако пропорциональность между активностью ферментов и их количеством может быть нарушена. В присутствии тормозящих или стимулирующих активность фермента веществ, на основе измерения активности могут быть сделаны ложные выводы о количестве фермента. Поэтому, измеряя активность фермента, нельзя судить о его количестве. Данные о количестве фермента можно получить лишь с учетом кинетики данной реакции.