

Действие ионизирующих излучений на белки
До 20 % поглощенной энергии связано с повреждением белков. Под действием ионизирующего излучения из молекулы белка выбивается электрон. Образуется дефектный участок, лишенный электрона, который мигрирует по полипептидной цепи за счет переброски соседних электронов до тех пор, пока не достигнет участка с повышенными электрон-донорными свойствами. В этом месте в боковых цепях аминокислот возникают свободные радикалы. Такие события происходят в результате прямого действия ионизирующих излучений. При косвенном действии образование свободных радикалов происходит при взаимодействии белковых молекул с продуктами радиолиза воды. Образование свободных радикалов влечет за собой изменения структуры белка:
разрыв водородных, гидрофобных, дисульфидных связей;
модификация аминокислот в цепи;
образование сшивок и агрегатов;
нарушение вторичной и третичной структуры белка.
Такие нарушения в структуре белка приводят к нарушению его функций (ферментативной, гормональной, рецепторной и др.).
Действие ионизирующих излучений на нуклеиновые кислоты
Около 7 % поглощенной дозы приходится на ядерную ДНК. Механизм повреждения сходен с повреждением белка: выбивание электрона, миграция дефектного участка по полинуклеотидной цепи (несколько сотен азотистых оснований) до участка с повышенными электрон-донорными свойствами. Такое место — чаще всего участок локализации тимина или цитозина, где и образуются свободные радикалы этих оснований. При косвенном действии ионизирующих излучений на нуклеиновые кислоты к образованию свободных радикалов приводит взаимодействие с продуктами радиолиза воды. Образование свободных радикалов приводит к нарушению структуры ДНК, в основе которого лежат следующие механизмы:
одно- и двунитевые разрывы;
модификация азотистых оснований;
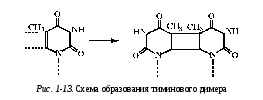
образование тиминовых димеров (рис. 1-13);
с
 шивки
ДНК–ДНК, ДНК-белок.
шивки
ДНК–ДНК, ДНК-белок.
При дозе 1 Гр в каждой клетке человека повреждаются около 5000 азотистых оснований, возникают примерно 1000 одиночных и от 10 до 100 двойных разрывов.
Определенное число одиночных разрывов образуется даже при малых дозах излучения, но они не приводят к поломкам молекулы ДНК, т. к. куски поврежденной молекулы прочно удерживаются на месте водородными связями с комплементарной нитью ДНК и хорошо поддаются восстановлению.
Репарация повреждений в ДНК является сложным процессом. Все механизмы репарации в клетке многократно продублированы и могут идти разными путями, находящимися под генетическим контролем. Некоторые виды репарации практически безошибочны. К ним относят фотореактивацию и эксцизионную репарацию коротких участков. В отличие от них, так называемая SOS-репарация часто бывает ошибочной, т. к. является попыткой восстановить структуру ДНК любой ценой при серьезных массивных повреждениях.
Репарация генетических повреждений обеспечивается десятками ферментов, многие из которых участвуют также в процессах репликации и рекомбинации. Известно 7 основных групп ферментов, участвующих в репарации:
нуклеозидазы (производят выщепление оснований по N-гликозидной связи с образованием так называемых АП-сайтов — апуриновых или апиримидиновых участков);
инсертазы (производят встраивание оснований в АП-сайты);
лиазы (производят расщепление пиримидиновых димеров);
эндонуклеазы (проводят инцизию — разрез ДНК возле повреждения);
экзонуклеазы (проводят эксцизию — удаление поврежденного участка);
ДНК-полимеразы (проводят синтез ДНК по комплементарной матрице);
ДНК-лигазы (производят сшивку нуклеотидов).
Рассмотрим основные типы репарации ДНК.
Прямая репарация. К ней относят несколько видов репарации:
Фотореверсия. Происходит за счет работы фотолиаз, причем начальный этап — образование фермент-субстратного комплекса — может идти и в темноте:
E + S ES + h E + P,
где Е – фермент (энзим), S – субстрат, P — продукт реакции.
Для работы фотолиазы требуется свет с длиной волны 350 нм. Этапы фотореверсии представлены ниже:
А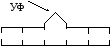 .
Повреждение ДНК с образованием димера
под действием УФ-излу-чения:
.
Повреждение ДНК с образованием димера
под действием УФ-излу-чения:
Б .
Образование фермент-субстратного
комплекса с эндонуклеазой:
.
Образование фермент-субстратного
комплекса с эндонуклеазой:
В. Восстановление структуры ДНК:

Восстановление одиночных разрывов (с участием ДНК-лигаз). Данный вид репарации весьма характерен при действии ионизирующих излучений, вызывающих образование большого числа однонитевых разрывов ДНК. Его этапы представлены ниже.
A .
Повреждение ДНК с образованием одиночного
разрыва:
.
Повреждение ДНК с образованием одиночного
разрыва:
Б. Образование фермент-субстратного комплекса с ДНК-лигазой:

В. Восстановление структуры ДНК:

Другие виды прямой репарации:
восстановление структуры азотистых оснований (удаление лишних метильных групп, восстановление разрывов циклических структур);
замена азотистых оснований (с участием ДНК-гликозидаз).
Репарация с использованием комплементарной цепи (эксцизионная репарация).
Существует несколько разновидностей эксцизионной репарации, наиболее изученная из которых — «темновая репарация». Основные этапы этого процесса одинаковы у различных видов организмов:
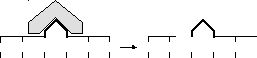
1 этап — incisio (разрезание) — эндонуклеаза «узнает» поврежденный участок и производит разрез:
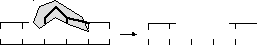 2
этап —
excisio (вычленение) — экзонуклеаза удаляет
поврежденный участок:
2
этап —
excisio (вычленение) — экзонуклеаза удаляет
поврежденный участок:
3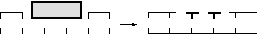 этап —
sintesis
— репаративный синтез с помощью
ДНК-полимеразы:
этап —
sintesis
— репаративный синтез с помощью
ДНК-полимеразы:
4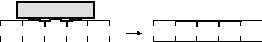 этап —
сшивка восстановленных участков
ДНК-лигазой:
этап —
сшивка восстановленных участков
ДНК-лигазой:
Репарация с использованием межмолекулярной информации. К этому типу репарации относят восстановление двойных разрывов и репарацию поперечных сшивок.
Восстановление двойных разрывов возможно только тогда, когда имеется копия генетической информации (например, при диплоидном наборе хромосом). В этом случае основа репарации — достаточно сложный процесс рекомбинации с реципрокным обменом нитей ДНК и последующим восстановлением повреждений. При этом образуются так называемые «структуры Холидея», которые в дальнейшем подвергаются разделению с образованием 2-х нормальных нитей ДНК.
Репарация поперечных сшивок внутри ДНК происходит по схеме «выщепление–рекомбинация–синтез». Ее этапы представлены ниже:
|
А. Выщепление |
Б. Рекомбинация |
В. Восстановление |
Г. Восстановление | ||||
|
участка 1’: |
Участков ДНК: |
структуры участков: |
структуры доменов: | ||||
|
1 |
2 2’ |
1 1’ |
2 2’ |
2 1’ |
1 2’ |
1 1’ |
2 2’ |
 1’
1’
Индуцибельная репарация. К этому виду репарации относят SOS-репарацию и постадаптационную репарацию.
SOS-репарация запускается в клетке при наличии сигнала бедствия — появления свободных фрагментов полинуклеотидной цепи, что указывает на серьезные повреждения ДНК. При этом клетка пытается восстановить структуру ДНК, невзирая на степень ее повреждения. Достигается это, в частности, снижением 3’-5’ — корректорской функции ДНК-полимеразы, что помогает быстро, но не всегда безошибочно, восстанавливать структуру.
Постадаптационная репарация впервые была описана при исследовании культуры лимфоцитов, которые, как известно, обладают повышенной чувствительностью к воздействию ионизирующих излучений. Оказалось, что после предварительного облучения культуры лимфоцитов при суммарной дозе около 30 сГр с низкой интенсивностью в течение 4 часов развивалась повышенная устойчивость к повреждению ДНК, длившаяся около 66 часов (3 клеточных цикла). Механизм этого явления еще до конца не изучен.
Таким образом, механизмы репарации генетических повреждений представляют собой сложную продублированную систему защиты генетической информации клетки и, следовательно, являются основой обеспечения надежности биологических систем. Именно поэтому большинство одиночных разрывов репарируются даже в летально облученных клетках и не являются причиной, определяющей гибель клетки. Однако нерепарированные одиночные разрывы могут в последующем привести к образованию двойных разрывов, плохо поддающихся восстановлению. Двойные разрывы могут возникнуть в результате единичного акта ионизации либо при совпадении одиночных разрывов на комплементарных нитях. Двойные разрывы опасны для клетки, т. к. они не всегда поддаются репарации и служат непосредственной причиной возникновения хромосомных аберраций. Основные виды хромосомных аберраций:
Фрагментация хромосом.
О
 бразование
хромосомных мостов, дицентриков,
кольцевых хромосом (рис. 1-14).
бразование
хромосомных мостов, дицентриков,
кольцевых хромосом (рис. 1-14).
Появление внутри- и межхромосомных обменов.
Часть аберраций (например, мосты) механически препятствуют делению клетки. Появление обменов, ацентрических фрагментов приводит к неравномерному разделению хромосом и утрате генетического материала, а это вызывает гибель клеток из-за недостатка метаболитов, синтез которых кодировался утраченной частью ДНК.