

Литический каскад и лизогенная репрессия
Некоторые фаги используют для выживания только одну линию поведения. Инфицируя чувствительную бактерию, они нарушают ее функции, с тем чтобы обеспечить образование большого числа фаговых частиц-потомков. В результате такой литической инфекции бактерия-хозяин погибает. В процессе типичного литического цикла фаговая ДНК (или РНК) проникает в клетку бактерии-хозяина, ее гены транскрибируются в установленном порядке, генетический материал фага реплицируется, и в результате образуются белковые компоненты фаговых частиц. В конечном счете бактериальная клетка разрушается (лизируется), освобождая зрелые частицы фагового потомства. Однако есть и такие фаги, для которых характерны две формы существования. Они способны воспроизводить себя путем такого же литического цикла, с помощью которого обеспечивается быстрое образование большого числа копий фаговых частиц. Однако развитие этих фагов может пойти и по другому пути. В этом случае фаговый геном находится в бактериальной клетке в латентной форме, называемой профагом. Такое состояние получило название лизогения. В лизогенных бактериях профаг интегрирован с бактериальным геномом и наследуется, как любая другая его часть, тем же способом, как и бактериальные гены. Благодаря наличию профага лизогенная бактерия обладает иммунитетом против инфицирования другими фаговыми частицами того же типа. Поэтому в бактериальном геноме обычно содержится только одна копия профага любого определенного типа. Переходы между лизогенным и литическим способами существования могут происходить в любом направлении. Если фаг, образованный в литическом цикле, проникает в клетку новой бактерии-хозяина, он может либо повторить литический цикл, либо перейти в лизогенное состояние. Выбор зависит от условий инфекции и генотипов фага и бактерии. Профаг может быть выведен из ли-зогенного состояния с помощью процесса, названного индукцией. В этом случае он исключается из бактериального генома и образует свободную фаговую ДНК, которая затем проходит через литический путь развития. Другой тип существования в бактериальной клетке представлен плазмидами. Это автономные элементы, геномы которых существуют в клетке как внехромосомные единицы. Плазмиды-это самореплицирующиеся кольцевые молекулы ДНК, которые сохраняются в клетке стабильно и в характерном для каждого типа плазмид числе копий; иными словами, число их остается постоянным из поколения в поколение. Некоторые из этих элементов также имеют альтернативные способы существования. Они могут находиться либо в автономном внехромосомном состоянии, либо могут быть интегрированы с бактериальной хромосомой ■ и находиться в ней как ее часть, подобно любой другой последовательности. Такие элементы получили название эписомы. (Следует отметить, что с тех пор, как было дано первоначальное определение, термины «плазмида» и «эписома» иногда свободно используются как взаимозаменяемые. Поэтому они не всегда точно соответствуют описанному здесь значению.)
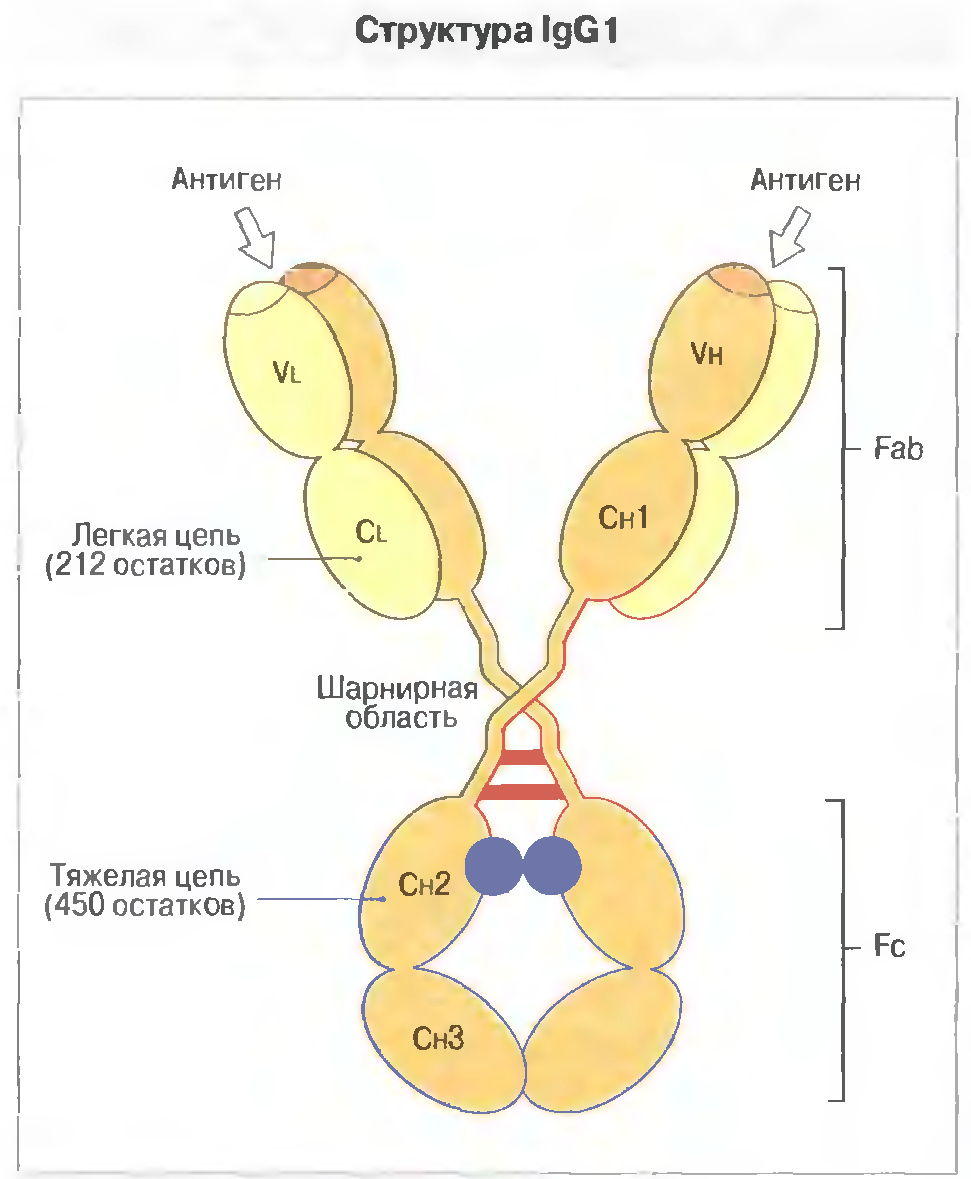
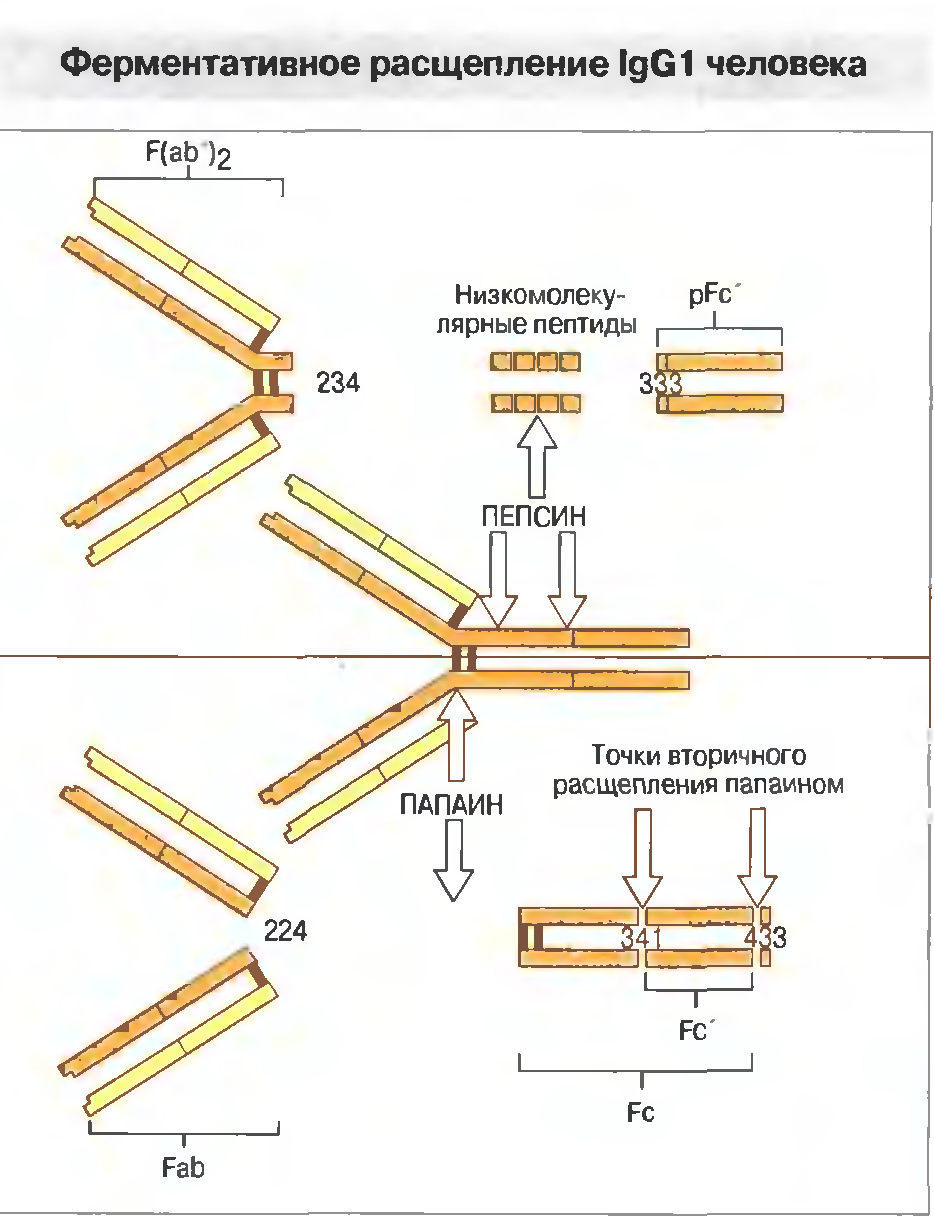
35. Молекулярна структура Імуноглобуліну G. Його характеристика.
Иммуноглобулин G ( IgG ) — монмер, основной класс AT (до 75% всех Ilg), защищающий организм от бактерий, вирусов и токсинов. После первичного контакта с Аг синтез IgM обычно сменяется образованием IgG.
Максимальные титры IgG при первичном ответе наблюдают на 6-8-е сутки. Обнаружение высоких титров IgG к Аг конкретного возбудителя указывает на то, что организм находится на стадии реконвалесценцш или конкретное заболевание перенесено недавно. В особо больших количествах IgG синтезируется при вторичном ответе.
IgG представлены 4 подклассами: IgG1, IgG2, IgG3 и IgG4; их относительное содержание (в %) составляет соответственно 66-70, 23, 7-8 и 2-4. IgG непосредственно участвуют в реакциях иммунного цитолиза, реакциях нейтрализации, а также усиливают фагоцитоз, действуя как опсонины и связывая рецепторы Fc-фрагмента в мембране фагоцитирующих клеток (в результате этого фагоциты эффективнее поглощают и лизируют микроорганизмы). Підкласи IgG відрізняються за своїми властивостями: здатністю до спонтанної агрегації, зв’язуванню комплементу. Їх біосинтез регулюється різними механізмами, за участю різних типів Т клітин.
Только IgG способны проникать через плаценту, что обеспечивает формирование у плода пассивного иммунитета. Міститься також у молозиві.
рганели руху бактеріальної клітини. Їх будова та механізм роботи.
По характеру движения подвижные бактерии разделяют на плавающие и скользящие(ползающие). Орган движения плавающих бактерий — жгутики; подвижность скользящих бактерий обеспечивают волнообразные сокращения тела.
Расположение жгутиков — характерный признак, имеющий таксономическое значение. Варианты расположения жгутиков приведены на рис. 4-1. У некоторых бактерий жгутики расположены по всей поверхности клеточной стенки (например, у бактерий рода Proteus), такие бактерии известны как перитрихи [от греч. peri-, вокруг, + trichos, волос]. Некоторые бактерии снабжены только одним толстым жгутиком (например, представители рода Vibrio), они известны как монотрихи. Политрихи — бактерии, имеющие одиночный по виду жгутик, образованный пучком из 2-50 жгутиков. Полярные жгутики прикреплены к одному или обоим концам бактерии. Монополярно-политрихиальное расположение жгутиков имеют лофотрихи [от греч. lophos, пучок, + trichos, волос], к ним, например, относят представителей рода Pseudomonas. Биполярно-политрихиальное жгутикование имеют амфитрихи [от греч. amphi-, двусторонний, + trichos, волос] (например, бактерии рода Spirillum).
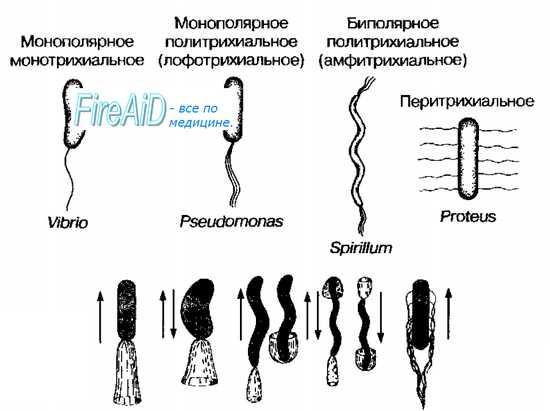
Рис. 4-1. Варианты расположения жгутиков (вверху) и движений бактерий (внизу).
Жгутик — спирально изогнутая полая нить, образованная субъединицами флагеллина. У разных бактерий толщина жгутиков варьирует от 12 до 18 нм, что составляет не более 1/10 диаметра жгутиков водорослей и простейших. Жгутики также различают по длине и диаметру витка. Место прикрепления жгутика к бактериальной клетке имеет сложное строение и состоит из базальной структуры и так называемого «крюка» (рис. 4-2). У грамположительных бактерий в состав базальной структуры входит одна пара, а у грамотрицательных бактерий — две пары колец. Кольца играют роль «приводного диска» и «подшипника». Вся конструкция выполняет функцию хемомеханического преобразователя (флагеллиновый мотор). У спирохет за движение ответственна особая органел-ла — осевая нить, состоящая из двух рядов бактериальных жгутиков, расположенных продольно внутри клетки.
Бактериальные жгутики совершают поступательные и вращательные движения, проталкивая бактерии через среду подобно корабельному винту. Они также могут изменять направление вращения и тянуть клетку подобно пропеллеру. Скорость обратного движения в четыре раза меньше скорости поступательного движения. Некоторые перитрихи могут перемещаться по поверхности агара, то есть плавающие бактерии способны к передвижению по поверхности твёрдых сред. В частности, Proteus vulgaris распространяется по поверхности агара, образуя тонкий налёт (напоминающий таковой при выдохе на холодное стекло), а неподвижные штаммы протея лишены такой способности. Это явление получило название «феномен роения», а наблюдение за ним легло в основу некоторых понятий бактериальной серодиагностики. Так, жгутиковые Аг называют Н-Аг [от нем. Hauch, выдох, налёт], а Аг клеточной поверхности — О-Аг [от нем. фпе Hauch, без налёта1.
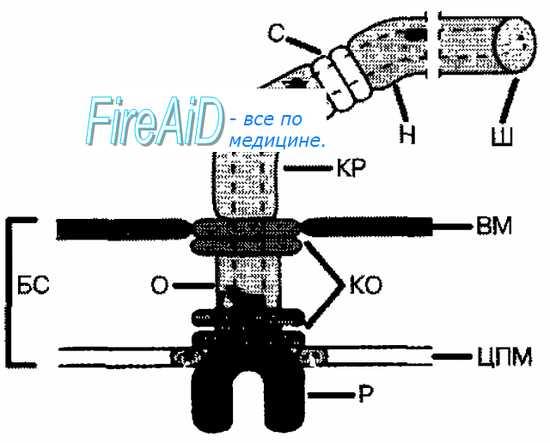
Рис. 4-2. Схема строения бактериального жгутика. БС — базальная структура, ВМ — внешняя мембрана, ЦПМ — цитоплазматическая мембрана, Р — ротор, О — ось, КО — кольцо жгутикового мотора, КР — крюк, С — цилиндрики-соединители, Н — нить жгутика, Ш — шапочка.
Способность бактерий к целенаправленному движению генетически обусловлена. Например, у Escherichia coli в регуляцию этого процесса вовлечено 3% генома (приблизительно 50 генов). Эти гены кодируют белки, образующие локомоторный аппарат, а также белки и ферменты, участвующие в преобразовании сигналов. Для жгутикового аппарата характерна периодическая изменчивость. Во многом этот процесс носит адаптивный характер и наиболее выражен у патогенных микроорганизмов. В частности, некоторые бактерии выработали систему вариабельности антигенных характеристик жгутиков, позволяющую им на какое-то время избегать направленных эффектов защитных иммунных механизмов.