

65. Основные законы наследуемости были описаны более века назад чешским монахом Грегором Менделем (1822-1884), преподававшим физику и естественную историю в средней школе г. Брюнна (г. Брно).
Мендель занимался селекционированием гороха, и именно гороху, научной удаче и строгости опытов Менделя мы обязаны открытием основных законов наследуемости: закона единообразия гибридов первого поколения, закона расщепления и закона независимого комбинирования.
Мендель экспериментировал с 22 разновидностями гороха, отличавшимися друг от друга по 7 признакам (цвет, текстура семян). Свою работу Мендель вел восемь лет, изучил 10 000 растений гороха. Все формы гороха, которые он исследовал, были представителями чистых линий; результаты скрещивания таких растений между собой всегда были одинаковы. Результаты работы Мендель привел в статье 1865 г., которая стала краеугольным камнем генетики. Трудно сказать, что заслуживает большего восхищения в нем и его работе – строгость проведения экспериментов, четкость изложения результатов, совершенное знание экспериментального материала или знание работ его предшественников.
Коллеги и современники Менделя не смогли оценить важности сделанных им выводов. По свидетельству А.Е. Гайсиновича, до конца XIX в. ее цитировали всего пять раз, и только один ученый – русский ботаник И.О. Шмальгаузен – оценил всю важность этой работы. Однако в начале XX столетия законы, открытые им, были переоткрыты практически одновременно и независимо друг от друга учеными К. Корренсом, Э. Чермаком и К. де Фризом. Значимость этих открытий сразу стала очевидна научному сообществу начала 1900-х годов; их признание было связано с определенными успехами цитологии и формированием гипотезы ядерной наследственности.
|
№66Роль наследственности и среды в развитии некоторых физических и психических признаков детей и подростков. |
|
|
Основные закономерности наследования признаков. Наследственность человека изучена в настоящее время в значительно меньшей степени, чем механизмы наследования признаков у растительных и животных организмов. Тем не менее уже сегодня получены интересные данные о наследственной обусловленности многих физических признаков человека. Современный уровень генетической науки позволяет также утверждать, что все основные закономерности наследования признаков и законы наследственности, выявленные в экспериментах с растениями и животными, являются справедливыми и для человека. Например, если молодой человек, у которого в обеих гомологичных хромосомах содержатся гены, обеспечивающие карий цвет глаз (гомозиготный по карему цвету), женится на голубоглазой девушке, у которой в хромосомах также содержатся только гены, обеспечивающие голубой цвет глаз, то их потомство будет наследовать цвет глаз по законам, открытым основателем генетики Г. Менделем еще в 60-е годы прошлого столетия в его опытах с растительными гибридами гороха. Наследование пола у человека и большинства животных также происходит по общим закономерностям и связано с распределением хромосом, имеющих Х- и К-образ-ную форму (половые хромосомы). В хромосомном наборе женского организма содержится две Х-хромосомы, а в мужском — одна хромосома X и одна У-хромосома (см. рис. 7). Число подобных примеров может быть бесконечно большим, и нет сомнения, что «генный портрет» человека (генотип) определяет в значительной степени многие его внешние свойства (фенотип). Ниже перечислены некоторые наследственные признаки человека, определяемые доминантными и рецессивными генами (по К- Вилли, 1974). Проявление генного влияния может осуществляться на различных этапах онтогенеза, но большинство фенотипи-ческих признаков определяется еще до рождения. Наконец, и само проявление генов не бывает фатальным, а зависит от факторов внешней среды. Например, тяжелое наследственное заболевание фенилкетонурия не развивается, если ребенок, содержащий в генотипе гены, вызывающие это заболевание, сразу же после рождения начинает получать определенную диету. Таким образом, генетика располагает убедительными фактами, доказывающими существование как строго обусловленных наследственностью^ признаков (например, уппы крови, гемофилия, цвет волос и глаз, черты лица и многие другие), так и признаков, определяемых в большой степени внешней средой (например, рост и масса человека, сила и ловкость его мышц, склонность к заболеваниям и др.). Следует отметить, что между генами и признаками не существует прямой связи: развитие одного признака может зависеть от влияния множества генов, а один ген может оказывать влияние на развитие многих признаков. Эта картина еще более усложняется постоянной коррекцией со стороны внешней среды. Следовательно, наел едет вен н ость _и с реaj^кaKjfcактотж^ развития всегда тесно взаимосвязаны. Нельзя представить себе развитие ребенка без корригирующих влияний среды, так же как оно невозможно без генетически предопределенной программы развития, представляющей синтез исторического опыта всех предшествующих поколений. Рассмотрим роль наследственности и среды в развитии некоторых физических признаков человека в процессе его пренатального и постнатального развития. Пренатальное развитие. Формирование органов и функциональных систем ребенка в процессе эмбриогенеза находится под контролем генотипа, но факторы внешней среды играют не последнюю роль. Для зародыша первичной средой является материнский организм. Здесь в специальном органе — матке — зародыш относительно хорошо защищен от многих вредных воздействий и получает через плаценту все необходимое ему для существования. Тем не менее, особенно на начальных этапах развития, многие факторы, влияющие на материнский организм, сказываются и на развитии зародыша (наиболее значительными являются: ионизирующее излучение, заболевания, перенесенные женщиной во время беременности, и многие химические вещества: алкоголь, никотин, антибиотики, гормональные препараты и др.). Следует отметить, что в пренатальном периоде человека есть критические периоды, когда развивающийся организм особо чувствителен к действию внешних факторов. Выделяют два таких периода. Первый включает начало пренатального развития, его первые три недели. В это время происходит закладка всех важнейших органов, и неблагоприятные воздействия в этот период чаще приводят к гибели зародыша. В течение второго критического периода (с 4-й по 7-ю неделю) происходит дальнейшее развитие всех органов, и вредные внешние воздействия в этот период могут привести к рождению ребенка с различными физическими дефектами (рис. 9). Важным является тот факт, что одинаковые физические врожденные дефекты могут быть связаны и с повреждением генетических структур, и с действием неблагоприятных внешних факторов в процессе эмбриогенеза. Это хорошее свидетельство того, что среда и наследственность в равной степени ответственны за нормальное развитие ребенка. Итак, к моменту рождения все органы человека и его физиологические системы, в том числе и нервная система, оказываются в общих чертах сформированными. Естественно, возникает вопрос: являются ли наследственно предопределенными те свойства нервной системы, которые лежат в основе психической деятельности человека, в основе его разума? Существуют ли готовые морфологические «заготовки сознания» так же, как они существуют для низшей нервной деятельности, связанной с регуляцией жизненных процессов нашего тела. Чтобы ответить на эти вопросы, остановимся на следующем, постнатальном этапе развития человека. Постнатальное развитие ребенка. Прежде всего рассмотрим основные методы, позволяющие изучать удельное значение среды и наследственности в постнатальном развитии детей и подростков. Проведение подобных экспериментов на растениях является простым и состоит в выделении двух групп организмов, идентичных по генотипу, и помещении этих групп в различные условия существования. Например, одну группу растений (контрольную) выращивают в обычных условиях, другую (экспериментальную) — в затемненном помещении. В результате подобных опытов можно сделать вывод, что образование зеленого хлорофилла растений зависит не только от наследственности, но и от факторов внешней среды (света), так как растения, выросшие в темноте, не будут содержать этот пигмент. Проведение аналогичных опытов на людях невозможно как с морально-этической стороны, так и с биологической. В мире нет двух людей с одинаковыми генотипами. Но... есть и исключения из правила. Это идентичные, или гомозиготные, близнецы (ИБ), имеющие не только поразительное внешнее сходство, но и почти одинаковые генные «портреты». Рождение идентичных и неидентичных близнецов (НБ) явление нередкое, в среднем из 100 беременностей одна завершается рождением более чем одного ребенка. Близнецы — превосходнейший «материал», подаренный ученым самой природой, поэтому и сам метод называют близнецовым. В чем его сущность? Допустим, нужно установить, какой фактор несет большую ответственность за физическое развитие и рост ребенка: наследственность или среда? Для этого выделяют близнецовые пары, проводят их морфологическое обследование и изучают образ жизни (обычно близнецы, особенно идентичные, имеют сходные интересы и близкие условия воспитания). На основании проведенных исследований устанавливают коэффициент сходства (конкордантность) между близнецами и делают заключение о роли наследственности или среды в развитии тех или иных качеств. Результаты подобных исследований идентичных и неидентичных близнецов по частоте встречаемости среди них некоторых заболеваний представлены в табл. К сожалению, педагоги проводят пока слабую работу по выявлению одаренных детей, что часто связано с недооценкой детской физиологической и психической индивидуальности, с незнанием элементарной биологии ребенка. Слабую работу по выявлению детской одаренности можно также объяснить еще низким уровнем исследований в этой области и отсутствием точных научных критериев, по которым педагогу нетрудно было бы обнаружить наследственные наклонности ребенка. Существует лишь тесная связь между одаренностью и высоким уровнем возбудимости нервной системы, признаком чего часто является резкая неуравновешенность (психопатичность) ребенка. К таким детям педагог должен относиться особо внимательно. Это необходимо прежде всего с гигиенической стороны, так как неверные действия педагога могут привести к развитию крайних черт характера в патологические. Необходимо это также и для своевременного выявления наследственных задатков и их оптимального развития. Важно помнить, что видимое отсутствие склонностей к учению, внешняя лен-ность и недисциплинированность еще не являются достаточными доказательствами отсутствия способностей. Из истории известно немало доказательств справедливости этого заключения. Создатель теории относительности А. Эйнштейн в детстве не проявлял открытых способностей и только в 9 лет смог пойти в подготовительную школу. Плохо учились в школе А. Гумбольдт (1769—1859) и Л. Пастер (1822—1895), выдающийся английский физик И. Ньютон слыл в школе лентяем и неспособным учеником. Можно полагать, что умелое обращение с ними родителей и педагогов создало в конце концов все необходимые условия для развития талантов. Тем более что важнейшим качеством гениальности является трудолюбие. Великий американский изобретатель Т. Эдисон писал: «Гений—это один процент вдохновения, а на девяносто девять процентов потение». Тот, кто не научится «потеть», никогда не сможет достичь больших успехов в любом виде человеческой деятельности, несмотря на самые выдающиеся наследственные задатки. Кто, как не педагог, должен в первую очередь воспитывать у ребенка трудолюбие — этот важнейший фактор прогрессивного развития личности, главнейшее условие формирования любых способностей. Таким образом, наследственность лишь дает «сырой материал», а среда осуществляет его основную «переработку». Человек, родившийся даже с самыми благоприятными задатками, но живущий в среде, препятствующей развитию его способностей, останется посредственностью, так что основная ответственность за развитие интеллектуальных возможностей ребенка ложится на воспитателей. Тем не менее роль среды не следует абсолютизировать, нельзя забывать, что «материал», поставляемый наследственностью, не всегда бывает качественным и даже самый талантливый скульптор не сможет из песка изваять скульптуру, точно так же, как глыба мрамора сама по себе не станет произведением искусства.
№67 - №68. Аутосомно-доминантное наследование — тип наследования, при котором одного мутантного аллеля, локализованного в аутосоме, достаточно, чтобы болезнь (или признак) могла быть выражена. Аутосомно-рецессивное наследование — тип наследования признака или болезни, при котором мутантный аллель, локализованный в аутосоме, должен быть унаследован от обоих родителей. |
№69. Наследование, сцепленное с полом — наследование какого-либо гена, находящегося вполовых хромосомах. Наследование признаков, проявляющихся только у особей одногопола, но не определяемых генами, находящимися в половых хромосомах,- называется наследованием, ограниченным полом.
Наследованием, сцепленным с X-хромосомой, называют наследование генов в случае, когда мужской пол гетерогаметен и характеризуется наличием Y-хромосомы (XY), а особи женского пола гомогаметны и имеют две X-хромосомы (XX). Таким типом наследования обладают все млекопитающие (в том числе человек), большинство насекомых и пресмыкающихся.
№70Голандрический тип наследования.Голандрическое наследование (holandricinheritance) — наследование признака, контролируемого геном, локализованным в негомологичной части Y-хромосомы. Голандрический ген (holandricgene) — ген, сцепленный с мужской половой Y-хромосомой. Признаки, определяемые Г.г, передаются только по мужской линии — от отца к сыну.
№71 При Х-сцепленном типе наследования мутантный ген расположен в X-хромосоме. Если при этом мутация обладает доминантным эффектом, то больными могут быть как мужчины, так и женщины. Однако от больного отца заболевание с вероятностью 100% передается только девочкам, но не мальчикам, получающим от отца Y-хромосому (рис. 5). Вероятность передачи доминантной Х-сцепленной мутации от больной матери детям составляет 50%. Болезнь с равной вероятностью может быть унаследована как дочерью, так и сыном.
К доминантному, сцепленному с Х-хромосомой, типу наследования относится известная детским врачам патология витамин Д-резистентный рахит (синонимы: гипофосфатемия, семейная-Х-сцепленная гипофосфатемия, фосфатдиабет). Диагноз этого тяжелейшего рахита, который не проходит под воздействием больших доз витамина Д, подтверждается наличием подобного заболевания у части родственников как мужского, так и женского пола.
Гораздо чаще Х-сцепленные заболевания наследуются по рецессивному типу. Отличительным свойством заболеваний с рецессивным Х-сцепленным типом наследования является то, что в семье болеют мужчины, а мутантный аллель они наследуют от своей, практически здоровой матери, гетерозиготной по мутантному аллелю. При составлении родословной у таких матерей нередко наблюдаются больные братья или дяди. Больные мужчины могут передавать свое заболевание только через поколение и только внукам (но не внучкам) через свою здоровую, но гетерозиготную дочь.
Таким образом, если проследить в родословной наследование по мужской линии рецессивного Х-сцепленного заболевания, то получится что-то вроде «хода шахматного коня».
Наиболее известными Х-сцепленными заболеваниями являются гемофилии А и В, а так же тяжелейшая патология мышечной системы - миодистрофияДюшенна Беккера. В основе развития гемофилии А лежат мутации гена, ответственного за синтез VIII фактора свертываемости крови, а при гемофилии В дефектным оказывается IX фактор свертываемости крови. Оба гена, кодирующие факторы VIII и IX, локализованы в длинном плече Х-хромосомы в областях q28 и q27.1-2, соответственно. Известно, что при гемофилии наблюдается нарушение свертываемости крови, и самые незначительные порезы могут привести больного без специальной гематологической помощи к летальному исходу. Отметим, что у женщин - носителей гена гемофилии (так называемых «кондукторов»), в отдельных случаях так же наблюдается склонность к кровотечениям, что выражается в обильных месячных и длительных кровотечениях во время родов. Это обстоятельство необходимо учитывать акушерам-гинекологам при работе с женщинами-носителями мутантных аллелей в любом из генов гемофилии А или В.
№72
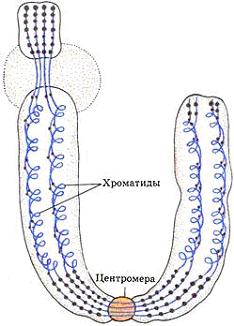
Строение и функция хромосом.Морфология
хромосом лучше всего видна в клетке на
стадии метафазы. Хромосома состоит из
двух палочкообразных телец - хроматид.
Обе хроматиды каждой хромосомы идентичны
друг другу по генному составу.
Хромосомы
дифференцированы по длине. Хромосомы
имеют центромеру или первичную перетяжку,
две теломеры и два плеча. На некоторых
хромосомах выделяют вторичные перетяжки
и спутники. Движение хромосомы определяет
Центромера, которая имеет сложное
строение.
ДНК центромеры отличается
характерной последовательностью
нуклеотидов и специфическими белками.
В зависимости от расположения центромеры
различают акроцентрические,
субметацентрические и метацентрические
хромосомы.
Как говорилось выше,
некоторые хромосомы имеют вторичные
перетяжки. Они, в отличие от первичной
перетяжки (центромеры), не служат местом
прикрепления нитей веретена и не играют
никакой роли в движении хромосом.
Некоторые вторичные перетяжки связаны
с образованием ядрышек, в этом случае
их называют ядрышковыми организаторами.
В ядрышковых организаторах расположены
гены, ответственные за синтез РНК.
Функция других вторичных перетяжек
еще не ясна.
У некоторых акроцентрических
хромосом есть спутники — участки,
соединенные с остальной частью хромосомы
тонкой нитью хроматина. Форма и размеры
спутника постоянны для данной хромосомы.
У человека спутники имеются у пяти пар
хромосом.
Концевые участки хромосом,
богатые структурным гетерохроматином,
называются теломерами. Теломеры
препятствуют слипанию концов хромосом
после редупликации и тем самым
способствуют сохранению их целостности.
Следовательно, теломеры ответственны
за существование хромосом как
индивидуальных образований.
Хромосомы,
имеющие одинаковый порядок генов,
называют гомологичными. Они имеют
одинаковое строение (длина, расположение
центромеры и т. д.). Негомологичные
хромосомы имеют разный генный набор и
разное строение.
 Исследование
тонкой структуры хромосом показало,
что они состоят из ДНК, белка и небольшого
количества РНК. Молекула ДНК несет
отрицательные заряды, распределенные
по всей длине, а присоединенные к ней
белки — гистоны заряжены положительно.
Этот комплекс ДНК с белком называют
хроматином. Хроматин может иметь разную
степень конденсации. Конденсированный
хроматин называют гетерохроматином,
деконденсированный хроматин —
эухроматином. Степень деконденсации
хроматина отражает его функциональное
состояние. Гетерохроматиновые участки
функционально менее активны, чем
эухроматиновые, в которых локализована
большая часть генов. Различают структурный
гетерохроматин, количество, которого
различается в разных хромосомах, но
располагается он постоянно в
околоцентромерных районах. Кроме
структурного гетерохроматина существует
факультативный гетерохроматин, который
появляется в хромосоме при
сверхспирализацииэухроматических
районов. Подтверждением существования
этого явления в хромосомах человека
служит факт генетической инактивации
одной Х-хромосомы в соматических клетках
женщины. Его суть заключается в том,
что существует эволюционно сформировавшийся
механизм инактивации второй дозы генов,
локализованных в Х-хромосоме, вследствие
чего, несмотря на разное число Х-хромосом
в мужском и женском организмах, число
функционирующих в них генов уравнено.
Максимально конденсирован хроматин
во время митотического деления клеток,
тогда его можно обнаружить в виде
плотных хромосом
Размеры молекул ДНК
хромосом огромны. Каждая хромосома
представлена одной молекулой ДНК. Они
могут достигать сотен микрометров и
даже сантиметров. Из хромосом человека
самая большая — первая; ее ДНК имеет
общую длину до 7 см. Суммарная длина
молекул ДНК всех хромосом одной клетки
человека составляет 170 см.
Несмотря
на гигантские размеры молекул ДНК, она
достаточно плотно упакована в хромосомах.
Такую специфическую укладку хромосомной
ДНК обеспечивают белки гистоны. Гистоны
располагаются по длине молекулы ДНК в
виде блоков. В один блок входит 8 молекул
гистонов, образуя нуклеосому (образование,
состоящее из нити ДНК, намотанной вокруг
октамера гистонов). Размер нуклеосомы
около 10 нм. Нуклеосомы имеют вид
нанизанных на нитку бусинок. Нуклеосомы
и соединяющие их участки ДНК плотно
упакованы в виде спирали, на каждый
виток такой спирали приходится шесть
нуклеосом. Так формируется структура
хромосомы.
Наследственная информация
организма строго упорядочена по
отдельным хромосомам. Каждый организм
характеризуется определенным набором
хромосом (число, размеры и структура),
который называется кариотипом. Кариотип
человека представлен двадцатью четырьмя
разными хромосомами (22 пары аутосом,
Х- и Y-хромосомы). Кариотип — это паспорт
вида. Анализ кариотипа позволяет
выявлять нарушения, которые могут
приводить к аномалиям развития,
наследственным болезням или гибели
плодов и эмбрионов на ранних стадиях
развития.
Длительное время полагали,
что кариотип человека состоит из 48
хромосом. Однако в начале 1956 г. было
опубликовано сообщение, согласно
которому число хромосом в кариотипе
человека равно 46.
Хромосомы человека
различаются по размеру, расположению
центромеры и вторичных перетяжек.
Впервые подразделение кариотипа на
группы было проведено в 1960 г. на
конференции в г. Денвере (США). В описание
кариотипа человека первоначально были
заложены два следующих принципа:
расположение хромосом по их длине;
группировка хромосом по расположению
центромеры (метацентрические,
субметацентрические, акроцентрические).
Точное
постоянство числа хромосом, их
индивидуальность и сложность строения
свидетельствуют о важности выполняемой
ими функции. Хромосомы выполняют функцию
основного генетического аппарата
клетки. В них в линейном порядке
расположены гены, каждый из которых
занимает строго определенное место
(локус) в хромосоме. В каждой хромосоме
много генов, но для нормального развития
организма необходим набор генов полного
хромосомного набора.
Исследование
тонкой структуры хромосом показало,
что они состоят из ДНК, белка и небольшого
количества РНК. Молекула ДНК несет
отрицательные заряды, распределенные
по всей длине, а присоединенные к ней
белки — гистоны заряжены положительно.
Этот комплекс ДНК с белком называют
хроматином. Хроматин может иметь разную
степень конденсации. Конденсированный
хроматин называют гетерохроматином,
деконденсированный хроматин —
эухроматином. Степень деконденсации
хроматина отражает его функциональное
состояние. Гетерохроматиновые участки
функционально менее активны, чем
эухроматиновые, в которых локализована
большая часть генов. Различают структурный
гетерохроматин, количество, которого
различается в разных хромосомах, но
располагается он постоянно в
околоцентромерных районах. Кроме
структурного гетерохроматина существует
факультативный гетерохроматин, который
появляется в хромосоме при
сверхспирализацииэухроматических
районов. Подтверждением существования
этого явления в хромосомах человека
служит факт генетической инактивации
одной Х-хромосомы в соматических клетках
женщины. Его суть заключается в том,
что существует эволюционно сформировавшийся
механизм инактивации второй дозы генов,
локализованных в Х-хромосоме, вследствие
чего, несмотря на разное число Х-хромосом
в мужском и женском организмах, число
функционирующих в них генов уравнено.
Максимально конденсирован хроматин
во время митотического деления клеток,
тогда его можно обнаружить в виде
плотных хромосом
Размеры молекул ДНК
хромосом огромны. Каждая хромосома
представлена одной молекулой ДНК. Они
могут достигать сотен микрометров и
даже сантиметров. Из хромосом человека
самая большая — первая; ее ДНК имеет
общую длину до 7 см. Суммарная длина
молекул ДНК всех хромосом одной клетки
человека составляет 170 см.
Несмотря
на гигантские размеры молекул ДНК, она
достаточно плотно упакована в хромосомах.
Такую специфическую укладку хромосомной
ДНК обеспечивают белки гистоны. Гистоны
располагаются по длине молекулы ДНК в
виде блоков. В один блок входит 8 молекул
гистонов, образуя нуклеосому (образование,
состоящее из нити ДНК, намотанной вокруг
октамера гистонов). Размер нуклеосомы
около 10 нм. Нуклеосомы имеют вид
нанизанных на нитку бусинок. Нуклеосомы
и соединяющие их участки ДНК плотно
упакованы в виде спирали, на каждый
виток такой спирали приходится шесть
нуклеосом. Так формируется структура
хромосомы.
Наследственная информация
организма строго упорядочена по
отдельным хромосомам. Каждый организм
характеризуется определенным набором
хромосом (число, размеры и структура),
который называется кариотипом. Кариотип
человека представлен двадцатью четырьмя
разными хромосомами (22 пары аутосом,
Х- и Y-хромосомы). Кариотип — это паспорт
вида. Анализ кариотипа позволяет
выявлять нарушения, которые могут
приводить к аномалиям развития,
наследственным болезням или гибели
плодов и эмбрионов на ранних стадиях
развития.
Длительное время полагали,
что кариотип человека состоит из 48
хромосом. Однако в начале 1956 г. было
опубликовано сообщение, согласно
которому число хромосом в кариотипе
человека равно 46.
Хромосомы человека
различаются по размеру, расположению
центромеры и вторичных перетяжек.
Впервые подразделение кариотипа на
группы было проведено в 1960 г. на
конференции в г. Денвере (США). В описание
кариотипа человека первоначально были
заложены два следующих принципа:
расположение хромосом по их длине;
группировка хромосом по расположению
центромеры (метацентрические,
субметацентрические, акроцентрические).
Точное
постоянство числа хромосом, их
индивидуальность и сложность строения
свидетельствуют о важности выполняемой
ими функции. Хромосомы выполняют функцию
основного генетического аппарата
клетки. В них в линейном порядке
расположены гены, каждый из которых
занимает строго определенное место
(локус) в хромосоме. В каждой хромосоме
много генов, но для нормального развития
организма необходим набор генов полного
хромосомного набора.
№73.В основе наследственности лежит способность всех живых организмов накапливать, хранить и передавать потомству наследственную информацию. Эта одна из важнейших качественных особенностей живой материи связана с нуклеиновыми кислотами — дезоксирибонуклеино-вой (ДНК) и рибонуклеиновой (РНК). Ведущее значение принадлежитДНК — самой длинной молекуле живых организмов, сосредоточенной в ядрах клеток и представляющей собой ее наследственный аппарат. Большая длина молекулы ДНК дает возможность «записать» на ней, как на телеграфной ленте, все основные свойства будущего организма и программу его развития. Такая «запись» осуществляется с помощью специального «нуклеинового языка», или «нуклеинового кода», сущностью которого является изменение порядка следования четырех химических соединений, входящих в состав ДНК Образно говоря, «нуклеиновый язык» состоит из четырех букв, из которых строятся отдельные слова и целые предложения «нуклеинового языка».
На такой нуклеиновой «ленте» можно выделить отдельные самостоятельные участки, включающие в себя описание программы развития одного признака. Их называют генами. Каждая молекула ДНК включает в себя сотни генов и представляет собой программу развития многих признаков и свойств организма. Объединяясь с особыми белками и некоторыми другими веществами, молекулы ДНК образуют в ядре специальные образования — так называемые хромосомы.
Организация генетического материала. Гены [от греч. genos, рождение] — единица наследственности, участок ДНК, занимающий специфическое место в хромосоме. С точки зрения генетики, ген — наследуемый фактор и неделимая единица генетического материала. Структурный ген (цистрон) — фрагмент ДНК, участвующий в образовании полипептидной цепи. В его состав входят лидерная последовательность, кодирующие фрагменты (экзоны), вставочные последовательности (нитроны) и концевая последовательность. Поскольку некоторые белки состоят более чем из одной субъединицы, формулировку «один ген — один фермент» применительно к гетеромультимерному (то есть состоящему из двух и более различных полипептидных субъединиц) белку следует трактовать как «один ген — одна полипептидная цепь».
Генотип — совокупность генов организма. Ещё в древности люди эмпирически использовали закономерности наследования. На основании этого опыта получила развитие селекция [от лат. selectio, выбирать] — наука о методах создания новых сортов растений и пород животных путём отбора и скрещивания. До недавнего времени генотип казался неприступным, не подвластным действиям человека. Открытие структуры генов позволило выделять их в изолированном виде, синтезировать биохимически и даже вводить в организм. Стало возможным воздействие на ген без его выделения из организма. Всё это создало предпосылки для манипулирования генотипом.
|
№73Наследственный аппарат клетки. В основе наследственности лежит способность всех живых организмов накапливать, |
|
№74 Строение нуклеиновых кислот.
Нуклеи́новаякисло́та (от лат.nucleus — ядро) — высокомолекулярное органическое соединение,биополимер(полинуклеотид), образованный остаткаминуклеотидов. Нуклеиновые кислотыДНКиРНКприсутствуют в клетках всех живых организмов и выполняют важнейшие функции по хранению, передаче и реализациинаследственной информации. Полимерные формы нуклеиновых кислот называют полинуклеотидами. Цепочки из нуклеотидов соединяются черезостаток фосфорной кислоты(фосфодиэфирная связь). Поскольку в нуклеотидах существует только два типа гетероциклических молекул, рибоза и дезоксирибоза, то и имеется лишь два вида нуклеиновых кислот —дезоксирибонуклеиновая(ДНК) ирибонуклеиновая(РНК).
Мономерные формы также встречаются в клетках и играют важную роль в процессах передачи сигналов или запасании энергии. Наиболее известный мономер РНК — АТФ, аденозинтрифосфорная кислота, важнейший аккумулятор энергии в клетке.
Одна из основных функций нуклеиновых кислот состоит в детерминации синтеза белков. Информация о структуре белков, закодированная в нуклеотидной последовательности ДНК, должна передаваться от одного поколения к другому, и поэтому необходимо ее безошибочное копирование, т.е. синтез точно такой же же молекулы ДНК (репликация).
№75Репликация ДНК-процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислотына матрице родительской молекулы ДНК. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение. Репликацию ДНК осуществляет сложный ферментный комплекс, состоящий из 15—20 различных белков, называемыйреплисомой(англ.replisome). Репликация ДНК — ключевое событие в ходеделения клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа:
инициация репликации
элонгация
терминация репликации.
Регуляция репликации осуществляется в основном на этапе инициации. Это достаточно легко осуществимо, потому что репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. Вгенометаких сайтов может быть как всего один, так и много. С понятием сайта инициации репликации тесно связано понятиерепликон. Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномыбактерий, как правило, представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации. Геномыэукариот(а также их отдельныехромосомы) состоят из большого числа самостоятельных репликонов, это значительно сокращает суммарное время репликации отдельной хромосомы. Молекулярные механизмы, которые контролируют количество актов инициации репликации в каждом сайте за один цикл деления клетки, называютсяконтролем копийности. В бактериальных клетках помимо хромосомной ДНК часто содержатсяплазмиды, которые представляют собой отдельные репликоны. У плазмид существуют свои механизмы контроля копийности: они могут обеспечивать синтез как всего одной копии плазмиды заклеточный цикл, так и тысяч копий.
Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка— место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более распространена двунаправленная репликация. Через некоторое время после начала репликации вэлектронный микроскопможно наблюдать репликационный глазок — участок хромосомы, где ДНК уже реплицирована, окруженный более протяженными участками нереплицированной ДНК.
В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500—5000 — у эукариот.
№76 Строение и виды хромосом.В ядре каждой соматической клетки организма человека содержится 46 хромосом. Набор хромосом каждого индивидуума, как нормальный, так и патологический, называется кариотипом.
Из 46 хромосом, составляющих хромосомный набор человека, 44 или 22 пары представляют аутосомные хромосомы, последняя пара — половые хромосомы. У женщин конституция половых хромосом в норме представлена двумя хромосомами X, а у мужчин — хромосомами X и У.
Во всех парах хромосом как аутосомных, так и половых одна из хромосом получена от отца, а вторая — от матери. Хромосомы одной пары называются гомологами, или гомологичными хромосомами. В половых клетках (сперматозоидах и яйцеклетках) содержится гаплоидный набор хромосом, т.е. 23 хромосомы.
 Сперматозоиды
делятся на два типа в зависимости от
того, содержат ли они хромосому X
или Y. Все яйцеклетки в норме содержат только
хромосому X.
Сперматозоиды
делятся на два типа в зависимости от
того, содержат ли они хромосому X
или Y. Все яйцеклетки в норме содержат только
хромосому X.
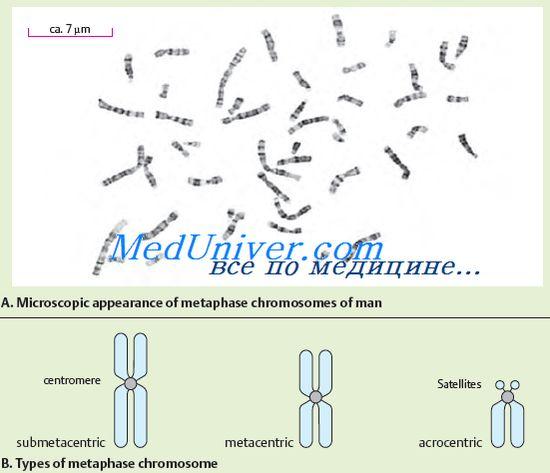
Хромосомы хорошо видны после специальной окраски во время деления клеток, когда хромосомы максимально спира-лизованы. При этом в каждой хромосоме выявляется перетяжка, которая называется центромерой. Центромера делит хромосому на короткое плечо (обозначается буквой «р») и длинное плечо (обозначается буквой «q»). Центромера определяет движение хромосомы во время клеточного деления. По положению центромеры хромосомы классифицируют на несколько групп. Если центромера располагается посредине хромосомы, то такая хромосома называется метацентриче-ской, если центромера располагается ближе к одному из концов хромосомы, то ее называют акроцентрической. Некоторые акроцентрические хромосомы имеют так называемые спутники, которые в неделящейся клетке формируют ядрышки. Ядрышки содержат многочисленные копии рРНК. Кроме того, различают субметацентрические хромосомы, когда центромера расположена не посредине хромосомы, а несколько сдвинута к одному из концов, но не столь значительно, как в акроцентрических хромосомах.
Концы каждого плеча хромосомы называют теломерами. Установлено, что теломеры играют важную роль в сохранении стабильности хромосом. В теломерах содержится большое число повторов последовательности нуклеотидов ТТАГГГ, так называемых тандемных повторов. В норме во время клеточного деления происходит уменьшение числа этих повторов в теломерах.
Однако каждый раз они достраиваются с помощью специального фермента, который называют теломеразой. Уменьшение активности этого фермента приводит к укорочению теломер, что, как полагают, является причиной гибели клеток, а в норме сопровождает старение.
№77.Денверская классификация хромосом. Классификация и номенклатура равномерно окрашенных хромосом человека впервые были приняты на международном совещании в 1960 году в г. Денвере, в дальнейшем несколько измененные и дополненные (Лондон, 1963 и Чикаго, 1966). Согласно Денверовской классификации все хромосомы человека разделены на 7 групп, расположенных в порядке уменьшения их длины и с учетом центриольного индекса (отношение длины короткого плеча к длине всей хромосомы, выраженное в процентах). Группы обозначаются буквами английского алфавита от А до G. Все пары хромосом принято нумеровать арабскими цифрами. Характеристика групп представлена в табл. 4. Предложенная классификация позволяла четко различать хромосомы, принадлежащие к различным группам. С 1960 года начинается бурное развитие клинической цитогенетики: в 1959 году Дж. Лежен открыл хромосомную природу синдрома Дауна; К. Форд, П. Джекобс и Дж. Стронг описали особенности кариотипа при синдромах Клайнфельтера и Тернера; в начале 70-х гг. была открыта хромосомная природа синдромов Эдвардса, Патау, синдрома «кошачьего крика»; описана хромосомная нестабильность при ряде наследственных синдромов и злокачественных заболеваниях. Вместе с тем применение метода получения равномерно окрашенных хромосом оказалось недостаточно эффективным для идентификации хромосом. Недостатком денверской классификации является то, что разграничение гомологичных пар внутри группы хромосом встречает зачастую непреодолимые трудности.
Таблица 4 ^ Группы хромосом и их характеристика
|
Группа |
№ хромосомы |
Расположение центромеры |
Центриольный индекс (%) |
Примечание |
|
А |
1 |
Самая большая метацентрическая |
48-49 |
На длинном плече может быть вторичная перетяжка |
|
|
2 |
Самая большая субметацентрическая |
38-40 |
|
|
|
3 |
Большая метацентрическая |
45-46 |
На 20% короче первой |
|
В |
4,5 |
Большая субметацентрическая |
24-30 |
|
|
С |
6-12 и Х-хромосома |
Средние субметацентрические |
27-35 |
На 9-ой часто вторичная перетяжка |
|
D |
13-15 |
Средние акроцентрические |
≈15 |
На всех вторичные перетяжки |
|
Е |
16 |
Маленькая метацентрическая |
40 |
В 10% случаев встречается вторичная перетяжка |
|
|
17 |
Маленькая субметацентрическая |
34 |
|
|
|
18 |
Маленькая субметацентрическая |
26 |
|
|
F |
19-20 |
Самые маленькие метацентрические |
36-46 |
|
|
G |
21-22 и Y-хромосома |
Самые маленькие акроцентрические |
13-33 |
На 21-й и 22-й вторичные перетяжки |
№78 Биосинтез белка в клетке. Процесс синтеза белка является очень сложным многоступенчатым процессом. Совершается он в специальных органеллах — рибосомах. В клетке содержится большое количество рибосом. Например, у кишечной палочки их около 20 000.
Каким образом происходит синтез белка в рибосомах?
Молекулы белков по существу представляют собой полипептидные цепочки, составленные из отдельных аминокислот. Но аминокислоты недостаточно активны, чтобы соединиться между собой самостоятельно. Поэтому, прежде чем соединиться друг с другом и образовать молекулу белка, аминокислоты должны активироваться. Эта активация происходит под действием особых ферментов. Причем каждая аминокислота имеет свой, специфически настроенный на нее фермент.
Источником энергии для этого (как и для многих процессов в клетке) служит аденозинтрифосфат (АТФ).
В результате активирования аминокислота становится более лабильной и под действием того же фермента связывается с т-РНК.
Важным является то, что каждой аминокислоте соответствует строго специфическая т-РНК. Она находит «свою» аминокислоту и переносит ее в рибосому. Поэтому такая РНК и получила название транспортной.
Следовательно, в рибосому поступают различные активированные аминокислоты, соединенные со своими т-РНК. Рибосома представляет собой как бы конвейер для сборки цепочки белка из поступающих в него различных аминокислот.
Оказывается, что одновременно с т-РНК, на которой «сидит» своя аминокислота, в рибосому поступает «сигнал» от ДНК, которая содержится в ядре. В соответствии с этим сигналом в рибосоме синтезируется тот или иной белок, тот или иной фермент (так как ферменты являются белками).
Направляющее влияние ДНК на синтез белка осуществляется не непосредственно, а с помощью особого посредника, той формы РНК, которая получила название матричной или информационной РНК (м-РНК или и-РНК).
Информационная РНК синтезируется в ядре иод влиянием ДНК, поэтому ее состав отражает состав ДНК. Молекула РНК представляет собой как бы слепок с формы ДНК.
Синтезированная и-РНК поступает в рибосому и как бы передает этой структуре план — в каком порядке должны соединяться друг с другом поступившие в рибосому активированные аминокислоты, чтобы синтезировался определенный белок. Иначе, генетическая информация, закодированная в ДНК, передается на и-РНК и далее на белок.
№79Мута́ция (лат. mutatio — изменение) — стойкое (то есть такое, которое может быть унаследовано потомками данной клеткиилиорганизма) изменениегенотипа, происходящее под влиянием внешней или внутренней среды. Термин предложенГуго де Фризом. Процесс возникновения мутаций получил названиемутагенеза.
Геномные: — полиплоидизация(образование организмов или клеток, геном которых представлен более чем двумя (3n, 4n, 6n и т. д.) наборами хромосом) ианеуплоидия(гетероплоидия) — изменение числа хромосом, не кратное гаплоидному набору (см. Инге-Вечтомов, 1989). В зависимости от происхождения хромосомных наборов среди полиплоидов различают аллополиплоидов, у которых имеются наборы хромосом, полученные при гибридизации от разных видов, и аутополиплоидов, у которых происходит увеличение числа наборов хромосом собственного генома, кратное n.
При хромосомных мутациях происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря (делеция) или удвоение части (дупликация) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах (инверсия), а также перенос части генетического материала с одной хромосомы на другую (транслокация) (крайний случай — объединение целых хромосом, т. н.Робертсоновскаятранслокация, которая является переходным вариантом от хромосомной мутации к геномной).
На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точечных мутациях. Поскольку в состав ДНК входятазотистые основаниятолько двух типов — пурины и пиримидины, все точечные мутации с заменой оснований разделяют на два класса: транзиции (замена пурина на пурин или пиримидина на пиримидин) и трансверсии (замена пурина на пиримидин или наоборот). Возможны четыре генетических последствия точковых мутаций: 1) сохранение смыслакодонаиз-за вырожденности генетического кода (синонимическая замена нуклеотида), 2) изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи (миссенс-мутация), 3) образование бессмысленного кодона с преждевременной терминацией (нонсенс-мутация). В генетическом коде имеются три бессмысленных кодона: амбер — UAG, охр — UAA и опал — UGA (в соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов — например амбер-мутация), 4) обратная замена (стоп-кодона на смысловой кодон).
По влиянию на экспрессию генов мутации разделяют на две категории: мутации типа замен пар оснований и типа сдвига рамки считывания (frameshift). Последние представляют собой делеции или вставки нуклеотидов, число которых не кратно трём, что связано с триплетностью генетического кода.
Первичную мутацию иногда называют прямой мутацией, а мутацию, восстанавливающую исходную структуру гена, — обратной мутацией, или реверсией. Возврат к исходному фенотипуу мутантного организма вследствие восстановления функции мутантного гена нередко происходит не за счет истинной реверсии, а вследствие мутации в другой части того же самого гена или даже другого неаллельного гена. В этом случае возвратную мутацию называют супрессорной. Генетические механизмы, благодаря которым происходит супрессия мутантного фенотипа, весьма разнообразны.
Почковые мутации (споры) — стойкие соматические мутации происходящие в клетках точек роста растений. Приводят к клоновой изменчивости. При вегетативном размножении сохраняются. Многие сортакультурных растений являются почковыми мутациями.
№80. Упаковка ДНК в хромосому.Хромосомы состоят из хроматина — смеси ДНК и белков в соотношении ~ 1:1. Именно благодаря взаимодействию с белками молекула ДНК имеет компактную структуру. Средняя длинна одной нити ДНК человека (если растянуть) 4 см. То есть в ядре каждой клетки содержится около 1 метра ДНК. Упаковка молекулы ДНК в хромосоме имеет три уровня (см. рисунок): 1.нуклеосомы, нуклеосомная нить 2. соленоидная спираль 3. петли Известная каждому своим видом двойная спираль ДНК «намотанная» вокруг комплекса из 4-х пар белковых молекул образует «бусину» - нуклеосому. Нуклеосомы, связанные между собой участками молекулы ДНК, составляют нить, которая в свою очередь закручена в форме соленоида (это по одной из существующих моделей), один виток — 6 нуклеосом. Эта структура на следующем уровне упаковки образует петли. В таком виде ДНК присутствует в ядре клетки на этапе роста клетки, синтеза её компонентов, в т. ч. удвоения ДНК. Однако на разных стадиях жизни клетки конденсация хроматина разная. В период роста клетки хроматин в ядре не конденсирован, компактность его расположения при этом отличается для разных участков хромосомы. Когда же происходит процесс деления клетки хроматин всё более уплотняется и петли, о которых говорилось выше, образуют «розетки», плотно уложенные одна к другой (тоже по одной из существующих моделей), по 18 петель на кольцо. В результате этого хроматин принимает вид конденсированных хромосом, так хорошо известный нам из учебников.
№81. Гаметогенез.(от Гаметы и греч. genesis — происхождение) процесс развития и формирования половых клеток — гамет (См. Гаметы). Г. мужских гамет (сперматозоидов, спермиев) называютСперматогенезом, женских гамет (яйцеклеток) —Оогенезом. У животных и растений Г. протекает различно, в зависимости от местаМейоза в жизненном цикле этих организмов.
У многоклеточных животных Г. происходит в специальных органах — половых железах, или гонадах (яичниках, семенниках, гермафродитных половых железах), и складывается из трёх основных этапов: 1) размножение первичных половых клеток — гаметогониев (сперматогониев и оогониев) путём ряда последовательных Митозов, 2) рост и созревание этих клеток, называют теперь гаметоцитами (сперматоцитами и ооцитами), которые, как и гаметогонии, обладают полным (большей частью диплоидным) набором хромосом (См.Хромосомы). В это время совершается основное событие Г. у животных — деление гаметоцитов путём мейоза, приводящее к редукции (уменьшению вдвое) числа хромосом в этих клетках и превращению их в гаплоидные клетки (см.Гаплоид) — сперматиды и оотиды; 3) формирование сперматозоидов (либо спермиев) и яйцеклеток; при этом яйцеклетки одеваются рядом зародышевых оболочек, а сперматозоиды приобретают жгутики, обеспечивающие их подвижность. У самок многих видов животных мейоз и формирование яйца завершаются после проникновения сперматозоида в цитоплазму ооцита, но до слияния ядер сперматозоида и яйцеклетки.
У растений Г. отделен от мейоза и начинается в гаплоидных клетках — в спорах (у высших растений — микроспоры и мегаспоры). Из спор развивается половое поколение растения — гаплоидный Гаметофит, в половых органах которого — гаметангиях (мужских — антеридиях (См.Антеридий), женских — архегониях (См.Архегоний)) путём митозов происходит Г. Исключение составляют голосеменные и покрытосеменные растения, у которых сперматогенез идёт непосредственно в прорастающей микроспоре — пыльцевой клетке. У всех низших и высших споровых растений Г. в антеридиях — это многократное деление клеток, в результате которого образуется большое число мелких подвижных сперматозоидов. Г. в архегониях — формирование одной, двух или нескольких яйцеклеток. У голосеменных и покрытосеменных растений мужской Г. состоит из деления (путём митоза) ядра пыльцевой клетки на генеративное и вегетативное и дальнейшего деления (также путём митоза) генеративного ядра на два спермия. Это деление происходит в прорастающей пыльцевой трубке. Женский Г. у покрытосеменных растений — обособление путём митоза одной яйцеклетки внутри 8-ядерного зародышевого мешка. Основное различие Г. у животных и растений: у животных он совмещает в себе превращение клеток из диплоидных в гаплоидные и формирование гаплоидных гамет; у растений Г. сводится к формированию гамет из гаплоидных клеток.
№82 Кариоти́п — совокупность признаков (число, размеры, форма и т. д.) полного набора хромосом, присущая клеткам данногобиологического вида(видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Кариотипом иногда также называют и визуальное представление полного хромосомного набора (кариограммы). Внешний вид хромосом существенно меняется в течениеклеточного цикла: в течениеинтерфазыхромосомы локализованы вядре, как правило, деспирализованы и труднодоступны для наблюдения, поэтому для определения кариотипа используются клетки в одной из стадий их деления —метафазе митоза. Для процедуры определения кариотипа могут быть использованы любые популяции делящихся клеток. Для определениячеловеческогокариотипа используют, как правило,лимфоцитыпериферической крови, переход которых от стадии покоя G0 кпролиферациипровоцируют добавлением митогенафитогемагглютинина. Для определения кариотипа могут быть использованы также клеткикостного мозгаили первичная культурафибробластовкожи. Для увеличения числа клеток на стадииметафазык культуре клеток незадолго перед фиксацией добавляютколхицинилинокадазол, которые блокируют образованиемикротрубочек, тем самым препятствуя расхождению хроматид к полюсам деления клетки и завершению митоза.
После фиксации препараты метафазных хромосом окрашивают и фотографируют; из микрофотографий формируют так называемый систематизированный кариотип — нумерованный набор пар гомологичных хромосом, изображения хромосом при этом ориентируются вертикально короткими плечами вверх, их нумерация производится в порядке убывания размеров, пара половых хромосом помещается в конец набора (см. Рис. 1).
Исторически первые недетализованные кариотипы, позволявшие проводить классификацию по морфологии хромосом, получали окраской по Романовскому — Гимзе, однако дальнейшая детализация структуры хромосом в кариотипах стала возможной с появлением методик дифференциального окрашивания хромосом. Наиболее часто используемой методикой вмедицинской генетикеявляется метод G-дифференциального окрашивания хромосом.
Сравнение комплексов поперечных меток в классической кариотипии или участков со специфичными спектральными характеристиками позволяет идентифицировать как гомологичные хромосомы, так и отдельные их участки, что позволяет детально определять хромосомные аберрации— внутри- и межхромосомные перестройки, сопровождающиеся нарушением порядка фрагментов хромосом (делеции,дупликации,инверсии,транслокации). Такой анализ имеет большое значение в медицинской практике, позволяя диагностировать ряд хромосомных заболеваний, вызванных как грубыми нарушениями кариотипов (нарушение числа хромосом), так и нарушением хромосомной структуры или множественностью клеточных кариотипов в организме (мозаицизмом).
№83Мейоз- редукционное деление, деления созревания, способ деления клеток, в результате которого происходит уменьшение (редукция) числа хромосом в два раза и одна диплоидная клетка (содержащая два набора хромосом) после двух быстро следующих друг за другом делений даёт начало 4 гаплоидным (содержащим по одному набору хромосом).
Биологическое значение М. заключается в поддержании постоянства Кариотипа в ряду поколений организмов данного вида и обеспечении возможности рекомбинации)хромосом и генов при половом процессе. М. — один из ключевых механизмов наследственности
Мейоз состоит из 2 последовательных делений с короткой интерфазой между ними.
Профаза I — профаза первого деления очень сложная и состоит из 5 стадий:
Лептотена или лептонема — упаковка хромосом, конденсация ДНК с образованием хромосом в виде тонких нитей (хромосомы укорачиваются).
Зиготена или зигонема — происходит конъюгация — соединение гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами и их дальнейшая компактизация.
Пахитена или пахинема — (самая длительная стадия) — в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них происходит кроссинговер — обмен участками между гомологичными хромосомами.
Диплотена или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток.
Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой.
К концу Профазы I центриоли мигрируют к полюсам клетки, формируются нити веретена деления, разрушаются ядерная мембрана и ядрышки
Метафаза I — бивалентные хромосомы выстраиваются вдоль экватора клетки.
Анафаза I — микротрубочки сокращаются, биваленты делятся и хромосомы расходятся к полюсам. Важно отметить, что, из-за конъюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе.
Телофаза I — хромосомы деспирализуются и появляется ядерная оболочка.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
Профаза II — происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления.
Метафаза II — унивалентные хромосомы (состоящие из двух хроматид каждая) располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку.
Анафаза II — униваленты делятся и хроматиды расходятся к полюсам.
Телофаза II — хромосомы деспирализуются и появляется ядерная оболочка.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и три так называемых редукционных тельца (абортивные дериваты первого и второго делений).
№84 Деление клеток – митоз
Если бы вы наблюдали под микроскопом за делением клеток, то увидели бы следующее. Во-первых, материал внутри ядра становится очень плотным. Можно даже различить дискретные структуры — хромосомы. Вы знаете, что в соматических (неполовых) клетках человека имеется 23 пары хромосом — по одной хромосоме из каждой пары от каждого родителя. Итого в соматических клетках человека 46 хромосом. Все 46 выстраиваются в экваториальной плоскости клетки. На этой стадии хорошо заметно, что каждая из 46 хромосом на самом деле состоит из двух хроматид, соединенных между собой в области, похожей на кнопку. Эта область была названа центромерой. С противоположных концов клетки расположено два плотных пятна (центриоли), каждое из которых образует нити веретена деления. Каждая нить присоединяется к хромосоме в области центромеры. К каждой паре хромосом присоединяются две нити — по одной от каждой центриоли. Затем мы бы увидели, что центриоли, подобно катушкам, наматывают нити веретена деления, в результате чего хромосомы расходятся к полюсам клетки. Затем делится содержимое клетки. Это удивительное зрелище.
События развиваются по следующему сценарию. Необходимо понимать, что для деления клетки должна быть решена очень важная задача — должно произойти удвоение (репликация) ДНК, ведь именно в ДНК закодирована вся информация о том, что и как должно происходить в клетке. Вспомните, что ДНК представляет собой двойную спираль. Снаружи спирали располагается сахарофосфатный остов, в котором сахара связаны друг с другом ковалентными связями через остаток фосфорной кислоты. Внутри спирали находятся органические азотистые основания, соединенные попарно водородными связями. Смысл копирования информации, хранящейся в одной хромосоме, и передачи этой информации новой клетке без изменений очень прост: необходимо раскрутить двойную спираль ДНК и разорвать водородные связи, удерживающие вместе пары азотистых оснований. С этого момента появляются две одинарные цепи с азотистыми основаниями, выступающими из сахарофосфатного остова. Вспомните, что каждое азотистое основание одной из цепей ДНК всегда образует связи со вполне определенным основанием другой цепи ДНК. Так, аденин связывается только с тимином, а гуанин — только с цитозином. В этом заключается принцип комплементарности азотистых оснований. Вспомните также, что нуклеотиды представляют собой молекулы, состоящие из органического основания и фосфорилированного сахара. Теоретически, если необходимые нуклеотиды присутствуют в нуклеоплазме, каждое азотистое основание каждой цепи ДНК подберет себе подходящую пару. Существует специфический и очень важный фермент, ДНК-полимераза, который делает эту реакцию возможной. Сахара вновь образуемой дочерней цепи ДНК соединяются друг с другом посредством фосфодиэфирных связей, в результате чего образуется сахарофосфатный остов — и вот оно свершилось! У вас есть две идентичные хромосомы там, где только что была одна (рис. 2.7). Конечно, на самом деле процесс репликации молекулы ДНК в клетке происходит намного сложнее и зависит от функционирования целого ряда ферментов, но об этом мы еще скажем ниже.А сейчас давайте более пристально рассмотрим процесс деления клетки. Вначале пары хромосом находятся в ядре в деспирализованном состоянии и неразличимы под микроскопом. О таких клетках говорят, что они находятся в интерфазе. В них генетический материал, известный как хроматин, представляет собой спутанную массу и не настолько компактен, насколько в то время, когда хромосомы различимы под микроскопом. Хроматин получил свое название из-за того, что он окрашивался в темный цвет специальными цветными красителями. Это произошло еще задолго до того, как люди узнали о его истинной функции. Как уже упоминалось, соматические клетки человека имеют 23 пары хромосом. Двадцать две пары представляют собой так называемые аутосомные хромосомы, одинаковые у мужчин и женщин. Двадцать третья пара представлена половыми хромосомами, которые различны у мужчин и женщин. Женщины имеют две большие хромосомы, названные X-хромосомами, а мужчины — одну большую Х-хромосому и одну маленькую Y-хромосому. У хромосом одной пары однотипное строение. Однако хромосомы, полученные от разных родителей, несут разную информацию, и в этом смысле хромосомные пары не идентичны.
Митоз, показанный на рис. 2.8, — универсальный способ деления любых эукариотических клеток. Митоз состоит из четырех основных четко различимых фаз: профазы, метафазы, анафазы, телофазы. Когда клетка приступает к делению, генетический материал каждой хромосомы уже удвоился. В профазе число хромосом соответствует 46, но каждая хромосома двойная, то есть состоит из двух сестринских хроматид.
Чтобы приступить к удвоению генетического материала, клетке предстоит решить трудную задачу — раскрутить спираль ДНК. Большинство из нас думают, что хромосома не так уж и длинна, потому что именно такими мы привыкли видеть их на всех рисунках. В действительности же в интерфазной клетке молекула ДНК находится в суперспирализованном состоянии, то есть скручена очень плотно и в определенном порядке. Показателен тот факт, что если бы молекула ДНК находилась внутри клетки в раскрученном состоянии, то ее длина составляла бы около 1 м! Только представьте себе, насколько сложной и упорядоченной должна быть упаковка такого большого количества генетического материала внутри микроскопической клетки. Отсюда становится понятным, почему в эукариотических клетках двойная спираль молекулы ДНК упакована в компактную структуру, образованную за счет взаимодействия с целым рядом белков, называемых гистонами. Таким образом, хромосомные белки упаковывают гигантскую молекулу в ядро объемом всего в несколько кубических микрон (рис. 2.9).Непосредственно перед началом процесса репликации молекула ДНК раскручивается ферментом хеликазой. Затем водородные связи разрываются, цепи расходятся, и на их матрице по принципу комплементарности образуются дочерние цепи.Деление клеток строго контролируется. Существуют клетки, которые делятся достаточно часто. К ним относятся клетки костного мозга. Другие клетки, например клетки кости, делятся главным образом во время активного роста органа или его повреждения. Наконец, существуют клетки, которые делятся очень редко или не делятся вообще. К такому типу относятся зрелые нервные клетки. Неконтролируемое деление и рост клеток является признаком их ракового перерождения. Процесс деления клеток требует регуляторов, которые называются факторами активации. Примерами таких фактории являются факторы роста. Изучение факторов роста одно из наиболее быстро развивающихся направлений на стыке современной биологии и медицины. Многие факторы в настоящее время выделены и частично охарактеризованы.
Факторы активации обычно работают, изменяя форму ДНК и позволяя ДНК-полимеразе, ответственной за репликацию ДНК, связаться с нею. ДНК-полимераза вначале находит специфический участок в молекуле ДНК. Этот участок, имеющий вид петли, необходим для связывания ДНК-полимеразы с ДНК. В свою очередь, он образуется при связывании фактора активации с ДНК. Участок молекулы ДНК, где происходит присоединение ДНК-полимеразы и начинается процесс репликации, называется точкой начала репликации. Следует также знать, что для инициации синтеза ДНК требуются короткие последовательности РНК, выполняющие функции «затравок» (праймеров). Связавшись с молекулой ДНК, ДНК-полимераза продвигается вдоль материнской цепи, достраивая дочернюю цепь по принципу комплементарности.
Одна из важнейших функций ДНК-полимеразы заключается в исправлении ошибок во вновь синтезированной цепи ДНК. Узнавая неправильные пары оснований, ДНК-полимераза возвращается на один шаг назад, неправильное основание удаляется из дочерней цепи, а правильное вставляется. В результате число ошибок при репликации ДНК составляет примерно одна ошибка на 10—100 млрд пар оснований. Однако, так как клетки делятся достаточно часто, ошибки неизбежно накапливаются. Многие из этих ошибок сказываются на жизнеспособности клеток. Если клетки, в которых произошла мутация, все же выживают, иммунная система разрушает их с помощью специальных, приспособленных для этих целей клеток.
Позвольте теперь вернуться к процессу деления клетки. В профазе митоза хромосомы состоят из двух хроматид, что является результатом репликации ДНК в интерфазе. Однако под микроскопом их регистрировать не удается, так как в профазе сестринские хроматиды тесно соприкасаются друг с другом, взаимно спирализуясь одна относительно другой. В прометафазе ядерная мембрана разрушается, и хромосомы начинают двигаться по направлению к экваториальной плоскости клетки. Этот процесс обеспечивает нормальное расхождение хромосом к полюсам клетки во время клеточного деления, в результате чего в каждую из вновь образующихся клеток попадает полный набор хромосом. Сестринские хроматиды одной хромосомы связаны друг с другом в точке, названной центромерой. В области центромеры расположен кинетохор — сложная белковая структура, связанная с ДНК центромерного участка. К этой зоне во время митоза прикрепляются нити веретена деления. Как вы уже наблюдали, на полюсах клетки расположены центриоли, число которых перед делением клетки также возрастает вдвое. Центриоли участвуют в формировании нитей веретена деления. Каждая нить веретена прикрепляется к одной хромосоме в области кинетохора. Во второй фазе митоза, метафазе, хромосомы, состоящие из двух хроматид, выстраиваются в экваториальной плоскости клетки.
Нити веретена деления растаскивают хромосомы, называемые с этого момента дочерними хромосомами, к противоположным концам клетки. Фаза, в течение которой хромосомы перемещаются в разные стороны, называется анафазой. Если все проходит по плану, центриоль делится, и сестринские хроматиды расходятся к полюсам клетки. Если хромосомы не выстраиваются в экваториальной плоскости клетки в должном порядке, процесс деления клетки может нарушиться.
После того как хромосомы разошлись к противоположным концам родительской клетки, клетка делится надвое в результате цитокинеза. Четвертая и заключительная стадия митоза называется телофазой. Клетки, размножающиеся путем митоза, как уже упоминалось, называются соматическими. К ним относятся все клетки нашего организма, за исключением клеток, которые впоследствии станут сперматозоидами или яйцеклетками. Клетки, которые вносят вклад в образование нового организма, называются половыми клетками. Половые клетки образуются в результате мейоза, который мы и рассмотрим в следующем разделе.
Митоз обычно проходит без какой-либо задержки. Однако в любой момент в организме человека происходят миллионы клеточных делений. Время от времени в клетках оказывается слишком много генетического материала, в то время как в других его может оказаться меньше нормы. Обычно такие клетки не выживают или удаляются иммунной системой.
№85 Наследственные болезни. Классификация наследственных болезней.Насле́дственныезаболева́ния — заболевания, возникновение и развитие которых связано с дефектами в программном аппаратеклеток, передаваемыми по наследству черезгаметы. Термин употребляется в отношенииполиэтиологических заболеваний, в отличие от более узкой группы —генные болезни.
В зависимости от вида первично поражённых клеток вьщелены следующие группы заболеваний: • болезни вследствие мутаций в половых клетках — «гаметические», т.е. собственно наследственные болезни (например, фенилкетонурия, гемофилии); эти болезни передаются по наследству; • болезни вследствие мутаций в соматических клетках — «соматические» (например, опухоли, некоторые болезни иммунной аутоагрессии). Эти заболевания не передаются по наследству; • болезни вследствие комбинации мутаций в половых и соматических клетках (например, семейная ретинобластома).
Отдельно рассматривают летальные, сублетальные и гипогенитальные заболевания: • летальные болезни приводят к гибели во время внутриутробного развития (например, моносомии по аутосомам, гаплоидии, большинство полиплоидии); • сублетальные заболевания приводят к гибели индивида до периода полового созревания (например, наследственные иммунодефициты типа агам-маглобулинемии швейцарского типа, синдром Луи-Бар, некоторые гемофилии); • гипогенитальные заболевания сочетаются с бесплодием (например, синдромы Шерешевского—Тернера, Кляйнфелтера).
В зависимости от роли наследственности и среды выделяют четыре группы заболеваний: • собственно наследственные болезни (развиваются вследствие мутаций); • экогенетические заболевания (развиваются при наличии мутации под влиянием специфического факторасреды); • болезни генетической предрасположенности (вклад генетических факторов очевиден); • болезни, вызываемые факторами внешней среды (строго говоря, эта группа болезней не относится к наследственным болезням, так как вклад генетических факторов в их возникновение, развитие и исход может быть крайне малым, сомнительным или неизвестным).
1
2