

Энергетическое значение ступенчатого транспорта электронов от окисляемых субстратов к молекулярному кислороду. Окислительное фосфорилирование в дыхательной цепи
Реакции прямого взаимодействия между кислородом и водородом в небиологических условиях сопровождаются взрывообразным выделением энергии в виде тепла и света. Многоступенчатый характер окислительных процессов в живой клетке обеспечивает постепенное освобождение энергии, которая может быть в дальнейшем использована в реакциях, связанных с выполнением различных видов биологической работы. Постепенное освобождение энергии уменьшает ее рассеивание и предохраняет клетку от разрушительного влияния тепловой энергии, которая при непосредственном взаимодействии окисляемого субстрата с кислородом освободилась бы одномоментно. Выработка тепла – это второстепенная функция процесса биологического окисления, основная функция биологического окисления заключается в обеспечении энергией процессов роста, анаболизма, транспорта веществ через мембраны, создания электрических потенциалов, механической работы и т.д.
Рис. 26.3. Строение
электронтранспортной (дыхательной)
цепи митохондрий. I
– NADH-дегидрогеназа;
II
– сукцинатдегидрогеназа; III
– цитохромы вс1,
IV
– цитохромоксидаза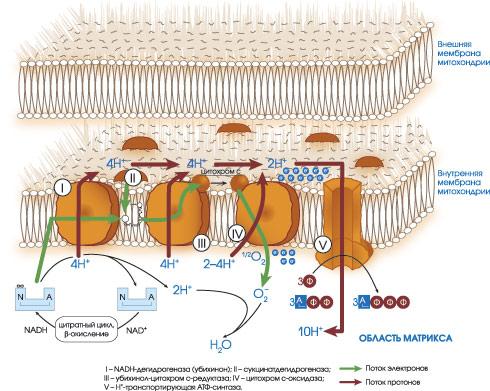
Синтез АТР из ADP и Н3РО4 за счет энергии, выделяющейся при тканевом дыхании, называется окислительным фосфорилированием. Сопряжение процессов окисления, служащих источником энергии, и фосфорилирования осуществляется в дыхательной цепи специальными механизмами, способными улавливать и переносить энергию потока высокоэнергетических электронов.
Митохондрии осуществляют важнейшую для клеточной биоэнергетики реакцию фосфорилирования АDP с образованием АТP за счет энергии окисления органических соединений, служащих субстратами окисления, молекулярным кислородом. Конечная стадия этого процесса – перенос электронов от восстановленных пиридиннуклеотидов и сукцината на молекулярный кислород – осуществляется по системе переносчиков электрона. При переносе электронов по дыхательной цепи происходит высвобождение энергии, величина которой (в электрон-вольтах) равна разности стандартных восстановительных потенциалов двух реагирующих редокс пар. Энергия одного моля иона в данной среде называется электрохимическим потенциалом. Разность электрохимических потенциалов протона между двумя водными фазами внутри и вне митохондрий описывается уравнением
![]()
где R ‒ газовая постоянная, T ‒ абсолютная температура, [H+]o и [H+]i ‒ концентрации ионов водорода вне и внутри матрикса, соответственно, F ‒ число Фарадея, ∆φ- разность потенциалов между окружающей средой и матриксом. П.Митчелл в качестве единицы энергии использовал электрон-вольты:
![]()
Суммарная энергия окислительно-восстановительной реакции, превращенная в разность электрохимических потенциалов ионов водорода, была названа П. Митчеллом протон-движущей силой (PMF ‒ proton motive force). Заменив натуральный логарифм на десятичный, легко найти величину протон-движущей силы, зная разность pH (pH) и разность потенциалов () между средой и матриксом при комнатной температуре; выраженная в миллиВ, она будет равна
PMF (мВ) = 60 (мВ) · pH +
В митохондриях основной вклад в эту сумму вносит мембранный потенциал, который в присутствии субстрата и кислорода составляет около 170-180 мВ. Созданная работой дыхательной цепи разность потенциалов может быть использована для синтеза АТР или переноса ионов в митохондрии. Синтез АТР осуществляется благодаря работе АТР-синтазы, которая представляет собой протонную АТРазу (H+-АТРазу).