

Регуляция биосинтеза хс
Скорость синтеза ХС регулируется по принципу отрицательной обратной связи. Основной пункт регуляции ‒ синтез мевалоновой кислоты ГМГ-СоА редуктазой. Холестерол подавляет ее синтез. При содержании 2-3 г холестерола в суточной пище человека синтез собственного ХС почти полностью прекращается.
Предполагается, что ХС или продукты его окисления в клетке могут угнетать непосредственно синтез редуктазы или индуцировать синтез ферментов, участвующих в ее деградации. При этом тормозится восстановление ГМГ-СоА в мевалоновую кислоту и синтез ХС в целом.
Скорость синтеза ГМГ-СоА редуктазы в печени подвергается четким суточным колебаниям. Максимум ее приходится на полночь, минимум ‒ на утренние часы. Активность ГМГ-СоА редуктазы возрастает при введении инсулина и тиреоидных гормонов. Угнетение активности фермента наблюдается при голодании, введении глюкагона и глюкокортикоидов.
Биосинтез желчных кислот
Желчные кислоты синтезируются в печени из холестерола. Синтез желчных кислот является доминирующим механизмом для удаления избытка холестерина. Сначала синтезируется из ХС холевая кислота; затем она подвергается АТР-зависимому активированию, превращаясь в холил-СоА (рис.24.8). Далее холил-СоА взаимодействует с таурином и глицином, образуя таурохолевую и гликохолевую кислоту.
Стадией, лимитирующей скорость образования желчных кислот, является реакция, катализируемая 7α-гидроксилазой. Холестерол в этой реакции требует участия электрон-транспортной системы, включающей цитохром Р-450 и NADPH-цитохром Р-450-редуктазу. Регулирующим фактором является концентрация желчных кислот, циркулирующих с желчью: чем выше их концентрация, тем ниже активность 7α-гидроксилазы, и наоборот.
Роль 7α-гидроксилазной реакции в образовании желчных кислот по значению сопоставима с ГМГ-СоА-редуктазной реакцией в биосинтезе холестерола.
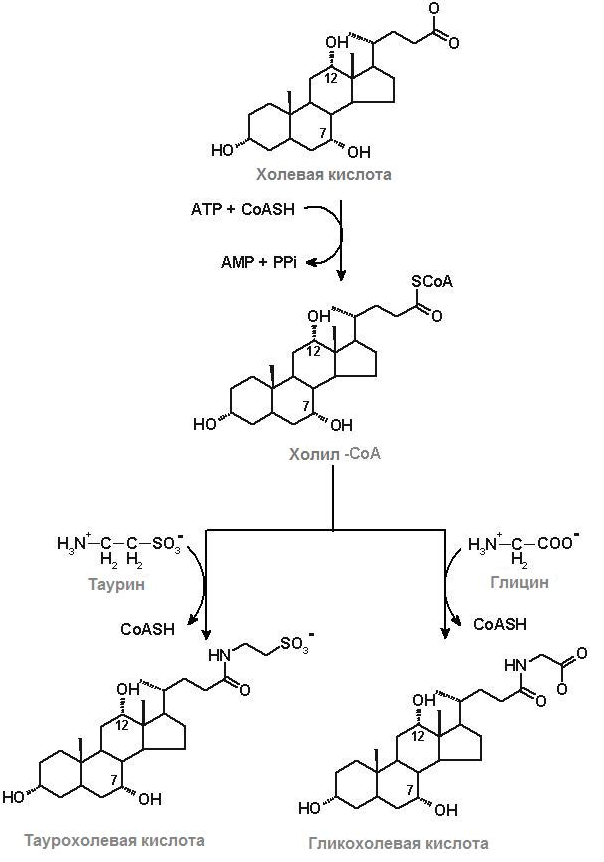
Рис.24.8. Образование таурохолевой и гликохолевой желчных кислот
Доказано, что активность этих ферментов изменяется параллельно, и т.к. как большая часть синтезируемого в печени холестерола (до 75%) идет на образование желчных кислот, часто бывает трудно определить, связано ли угнетение их образования с подавлением ГМГ-СоА-редуктазы или 7α-гидроксилазы.
Лекция 25
БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ
Роль высокоэнергетических фосфатов в биоэнергетике.
Биологическая роль АТР. Свободная энергия
гидролиза АТФ и других органических фосфатов
Для описания состояния химической системы используют понятие свободной энергии Гиббса, которое ввели Гиббс и Гельмгольц, объединив 1-й и 2-й законы термодинамики: ΔG = ΔЕ – TΔS. Определение «свободная» означает свободу использовать энергию для совершения полезной работы. Свободная энергия ΔG количественно характеризует потенциальную способность вещества претерпевать химические и физические превращения. Химическая реакция протекает лишь в том случае, если ∆G<0, т.е. в условиях, когда свободная энергия продуктов реакции меньше, чем свободная энергия исходных веществ. В химических процессах ΔG зависит не только от характера реакции, но и от тех условий, в которых она протекает.
Все содержащиеся в клетке фосфорилированные соединения разделяют на две группы: высокоэнергетические и низкоэнергетические (в зависимости от величины ΔG0ʹ их гидролиза). Высокоэнергетические фосфаты выполняют в клетке функцию аккумулирования биологической энергии и ее последующего использования для выполнения клеточных функций. К таким фосфатам относятся креатинфосфат (ΔG0ʹ= -14,80 ккал), фосфоенолпируват (ΔG0ʹ= -11,80 ккал), карбамоилфосфат. Изменение стандартной свободной энергии гидролиза АТР при t=37С и рН 7,0 в присутствии избытка Mg2+ составляет -7,3 ккал/моль.
АТР в термодинамической шкале занимает промежуточное положение. Этим объясняется уникальность молекулы АТР и ее биологическая роль посредника при переносе фосфатных групп от высокоэнергетических фосфорилированных соединений к акцепторным молекулам. Запас АТР в клетке может обеспечить энергией работу клетки лишь на несколько секунд. Цикл АТР-ADP работает постоянно и производит такое количество АТР, которое было израсходовано клеткой. За сутки в организме человека образуется и распадается около 60 кг АТР. Энергия химических связей АТР используется в организме для совершения полезной работы. На всех этапах превращения энергии, в том числе и при гидролизе АТР, часть энергии выделяется в виде тепла.
Высвобождение большого количества энергии при гидролизе АТР объясняется структурными особенностями этой молекулы: 1) молекулы АТР при рН=7,0 имеют в среднем около 3,8 отрицательных заряда. Эти заряды располагаются близко друг от друга и потому между ними – сильное отталкивание. При гидролитическом отщеплении концевой фосфатной группы сила отталкивания уменьшается. Образовавшиеся продукты (анионы НРО42- и ADP3-) не стремятся к объединению, потому что их сближению препятствует отталкивание одноименных зарядов. 2) продукты гидролиза, НРО42- и ADP3- стабилизируются за счет сопряжения. Электроны, окружающие атомы фосфора и кислорода концевой фосфатной связи АТР, конкурируют за энергетически выгодные орбитали. Наличие такой конкуренции не позволяет всем электронам концевой пирофосфатной связи занять столь низкие энергетические уровни, какие они способны занять в отдаленных друг от друга ионах НРО42- и ADP3-.