

Спиртовое и молочнокислое брожение
Спиртовое и молочнокислое брожение – основные источники обеспечения энергией некоторых микроорганизмов в анаэробных условиях.
Спиртовое брожение осуществляется так называемыми дрожжеподобными организмами, а также некоторыми плесневыми грибками. Суммарную реакцию спиртового брожения можно изобразить следующим образом:
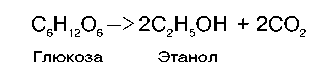
Механизм процесса спиртового брожения чрезвычайно близок к гликолизу. Расхождение начинается лишь после этапа образования пирувата. При гликолизе пируват при участии фермента ЛДГ и кофермента NADH восстанавливается в лактат. При спиртовом брожении этот конечный этап заменен двумя другими ферментативными реакциями: пируватдекарбоксилазной и алкогольдегидрогеназной.
В дрожжевых клетках (спиртовое брожение) пируват вначале подвергается декарбоксилированию, в результате чего образуется ацетальдегид. Данная необратимая реакция катализируется ферментом пируватдекарбоксилазой, который требует наличия ионов Mg2+ и кофермента (TРP):

Образовавшийся ацетальдегид присоединяет к себе водород, отщепляемый от NADH, восстанавливаясь при этом в этанол. Реакция катализируется ферментом алкогольдегидрогеназой:

Таким образом, конечными продуктами спиртового брожения являются этанол и СО2, а не молочная кислота, как при гликолизе.
Процесс молочнокислого брожения имеет большое сходство со спиртовым брожением. Отличие заключается лишь в том, что при молочнокислом брожении пировиноградная кислота не декарбоксилируется, а, как и при гликолизе в животных тканях, восстанавливается при участии ЛДГ за счет водорода NADH.
Известны две группы молочнокислых бактерий. Бактерии одной группы в процессе брожения углеводов образуют только молочную кислоту, а бактерии другой из каждой молекулы глюкозы «производят» по одной молекуле молочной кислоты, этанола и СО2.
Существуют и другие виды брожения, конечными продуктами которых могут являться пропионовая, масляная и янтарная кислоты, а также другие соединения.
Лекция 18
Аэробный катаболизм углеводов (часть 1)
Аэробный метаболизм пирувата. Митохондрии: структура
И энергетические функции
Клетки, недостаточно снабжаемые кислородом, могут частично или полностью существовать за счет энергии гликолиза. Однако большинство животных и растительных клеток в норме находится в аэробных условиях и свое органическое «топливо» окисляет полностью (до СО2 и Н2О). В этих условиях пируват, образовавшийся при расщеплении глюкозы, не восстанавливается до лактата, а постепенно окисляется до СО2 и Н2О в аэробной стадии катаболизма, при этом первоначально происходит окислительное декарбоксилирование пирувата с образованием ацетил-CоА.
Окислительное декарбоксилирование пирувата. Строение
мультиферментного пируватдегидрогеназного комплекса. Суммарное уравнение и энергетический баланс окислительного
декарбоксилирования пирувата. Регуляция активности
пируватдегидрогеназного комплекса: ковалентная модификация,
аллостерический механизм
Окисление пирувата до ацетил-CоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
На первой стадии этого процесса пируват (рис. 18.1) теряет свою карбоксильную группу в результате взаимодействия с тиаминпирофосфатом (TPP) в составе активного центра фермента пируватдегидрогеназы (E1). На второй стадии оксиэтильная группа комплекса E1–TPP–СНОН–СН3 окисляется с образованием ацетильной группы, которая одновременно переносится на амид липоевой кислоты (кофермент), связанной с ферментом дигидроли-поилацетилтрансферазой (Е2). Этот фермент катализирует третью стадию – перенос ацетильной группы на коэнзим CоА (HSCoA) с образованием конечного продукта (ацетил-CоА), который является высокоэнергетическим (макроэргическим) соединением.
На четвертой стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамида-Е2. При участии фермента дигидролипоилдегидрогеназы (Е3) осуществляется перенос атомов водорода от восстановленных сульфгидрильных групп дигидролипоамида на FAD, который выполняет роль простетической группы данного фермента и прочно с ним связан. На пятой стадии восстановленный FADН2 дигидро-липоилдегидрогеназы передает водород на кофермент NAD+ (с образованием NADН + Н+).
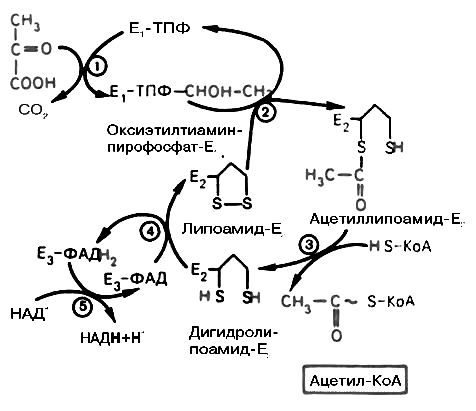
Рис. 18.1. Механизм действия пируватдегидрогеназного комплекса. Е1 – пируватдегидрогеназа; Е2 – дигидролипоилацетилтрансфераза; Е3 – дигидролипоилдегидрогеназа (цифры в кружках обозначают стадии процесса)
Процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий. В нем (в составе сложного мультиферментного комплекса) принимают участие 3 фермента (пируватдегидрогеназа, ди-гидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 кофер-ментов (TРP, амид липоевой кислоты, коэнзим А, FAD и NAD), из которых три относительно прочно связаны с ферментами (ТPP-E1, липоамид-Е2 и FAD-Е3), а два – легко диссоциируют (HS-CoA и NAD).
Все эти ферменты, имеющие субъединичное строение, и коферменты организованы в единый комплекс. Поэтому промежуточные продукты способны быстро взаимодействовать. Доказано, что полипептидные цепи субъединиц дигидролипоил-ацетилтрансферазы составляют как бы ядро комплекса, вокруг которого расположены пируватдегидрогеназа и дигидролипоилдегидрогеназа. Принято считать, что нативный ферментный комплекс образуется путем самосборки.
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:
![]()
Реакция сопровождается значительным уменьшением стандартной свободной энергии и практически необратима (∆G0′= ‒ 40 кДж/моль).
Образовавшийся в процессе окислительного декарбоксилирования аце-тил-CоА подвергается дальнейшему окислению с образованием СО2 и Н2О. Полное окисление ацетил-CоА происходит в цикле трикарбоновых кислот (цикл Кребса). Этот процесс, так же как окислительное декарбо-ксилирование пирувата, происходит в митохондриях клеток.