

Олигосахариды
Олигосахариды – сложные углеводы, молекулы состоят из небольшого числа (от 2 до 10) остатков моносахаридов. Олигосахариды, состоящие из трех, четырех и пяти остатков моносахаридов, соответственно называют так: три-, тетра-, и пентасахариды. Среди всех олигосахаридов наибольшее значение имеют дисахариды (биозы). Они имеют общую формулу С12Н22О11. Дисахариды содержат два моносахаридных остатка, соединенных О-гликозидной связью, которая формируется тогда, когда гидроксильная группа одной молекулы моносахарида реагирует с гидроксильной группой другой. Связь, образованная между остатками моносахаридов, может быть достаточно легко гидролизована кислотами, но является достаточно устойчивой по отношению к щелочному гидролизу:
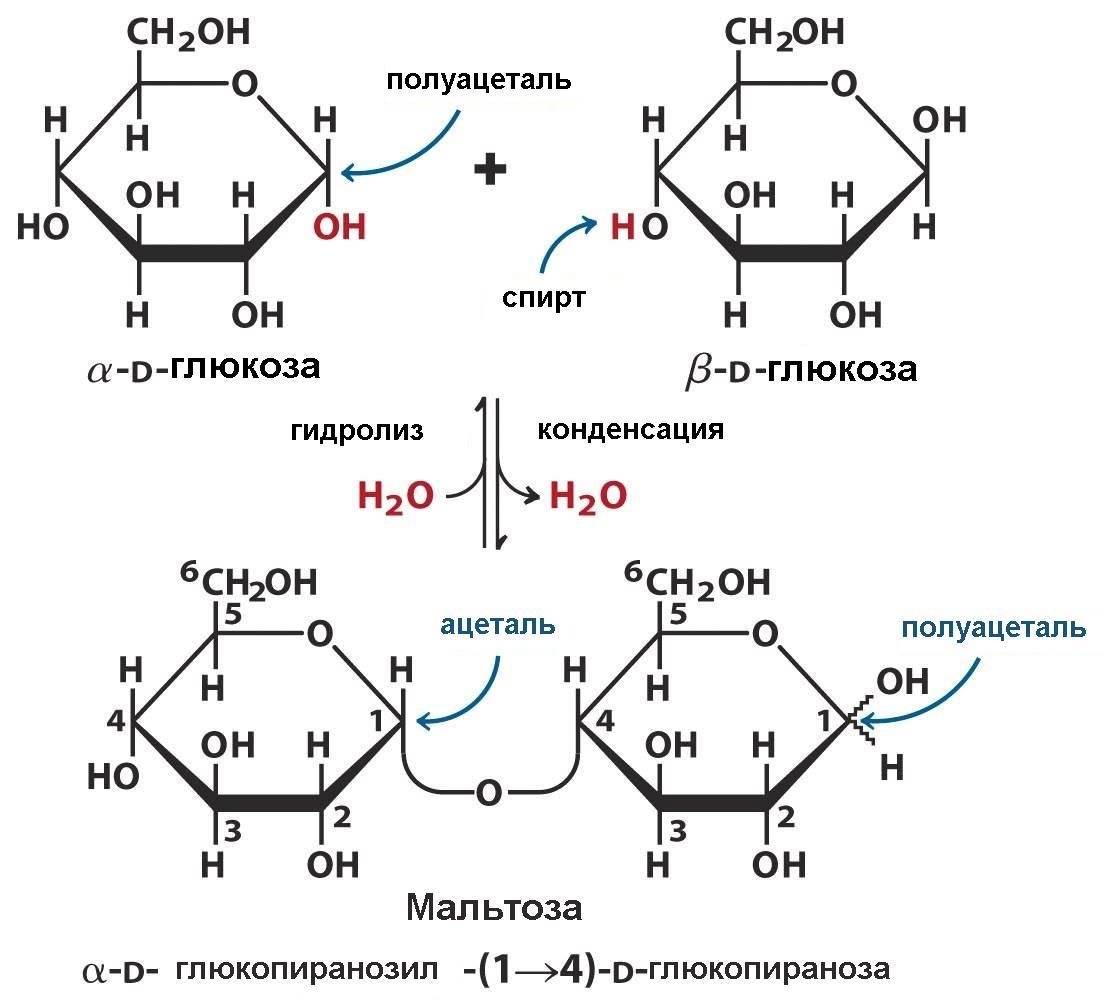
Известно, что альдозы, находящиеся в открытой, линейной форме, и обладающие, таким образом, свободной альдегидной группой, отличаются, как и другие альдегиды, восстанавливающими свойствами. Например, они могут давать «реакцию серебряного зеркала». Оказывается, что и некоторые дисахариды также обладают восстанавливающими свойствами. Это такие дисахариды, у которых сохраняется полуацетальный гидроксил одного из остатков моносахаридов. Примером восстанавливающих дисахаридов является мальтоза.
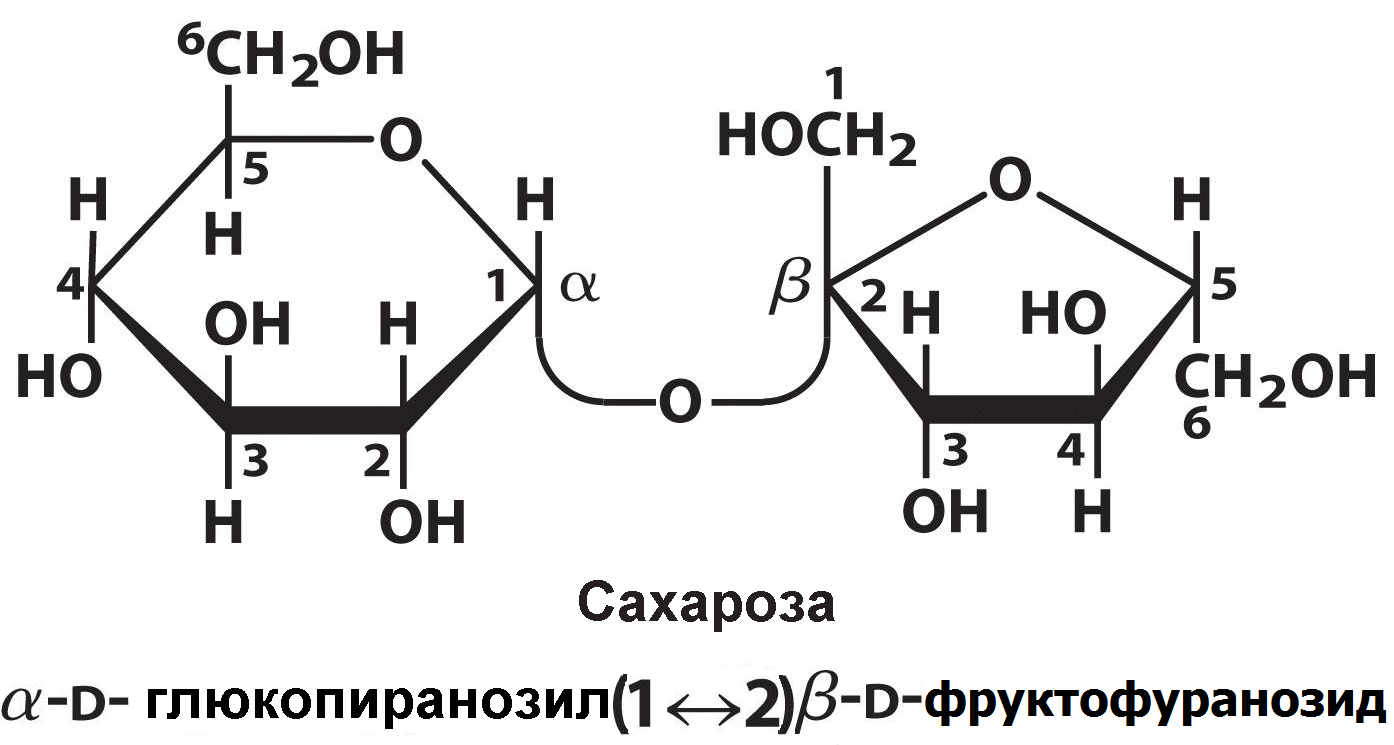
В противоположность ей, в молекуле сахарозы нет свободного гликозидного гидроксила, и поэтому она не обладает восстанавливающими свойствами:
Номенклатура дисахаридов: восстанавливающие дисахариды называются гликозилальдозами или кетозами, а невосстанавливающие – гликозилальдозидами или кетозидами. Краткое написание структуры дисахаридов предусматривает написание моносахаридных остатков подряд, начиная с невосстанавливающего конца, с использованием буквенных обозначений моносахаридных остатков, указанием конформации D и L, цифрами атомы, через которые осуществляется связь, характер кольца – пираноза или фураноза, стрелкой – направление связи.
Например, сахароза: α-D-Glcp (1→2)-β-D-Fruf; мальтоза: α-D-Glcp (1→4) α-D-Glcp; лактоза: β-D-Galp (1→4) α-D-Glcp; изомальтоза: α-D-Glcp (1 →6) α-D-Glcp. В повседневной практике чаще употребляются рабочие, тривиальные названия, многие из которых указывают или на происхождение данного дисахарида, или на его свойства. Тривиальные названия формируются с добавлением окончания – оза.
Рациональные названия дисахаридов: сахароза – α-D-глюкопиранозил (1→2)-β-D-фруктофуранозид; лактоза ‒ β-D-галактопиранозил (1→4)-α-D-глюкопираноза.
Отдельные представители дисахаридов
Мальтоза, или солодовый сахар (от лат. maltum – солод), является продуктом неполного гидролиза крахмала. Образуется под влиянием ферментов, содержащихся в солоде.
Изомальтоза – входит в состав амилопектиновой фракции крахмала и гликогена, связь α (1→6).
Целлобиоза – повторяющееся звено целлюлозы, связь β(1→4); широко распространена в растительном мире.
Лактоза (молочный сахар) в значительных количествах находится в молоке, имеет важное значение для растущих организмов, как животных, так и человека. В коровьем молоке содержится до 4,5% лактозы, в женском молоке – до 7,5%. При гидролизе, например в кишечнике во время переваривания пищи, лактоза распадается на α-D-глюкозу и β-D-галактозу.
Сахароза (тростниковый сахар) служит растворимым резервным сахаридом растений. В больших количествах содержится в сахарной свекле, сахарном тростнике и кленовом соке, из которых ее получают в промышленных масштабах. Сахароза является наиболее известным дисахаридом, т.к. чрезвычайно широко используется в пищевой промышленности и в домашнем питании.
Гидролиз сахарозы ([α]D20 = +66,5о) с образованием глюкозы ([α]D20 = + 52,5о) и фруктозы ([α]D20 = ‒ 92о) часто называют инверсией, поскольку он сопровождается изменением знака оптического вращения, правовращающая сахароза превращается в левовращающую смесь эквимолярных количеств глюкозы и фруктозы. Эту смесь часто называют инвертным сахаром.
Трегалоза – грибной сахар, состоит из двух остатков α D-Glc, соединенных α (1→1)-гликозидной связью. Встречается в грибах, спорынье, водорослях и некоторых других растениях. Является главным углеводом гемолимфы многих насекомых.