

Витамин в6 (пиридоксин, пиридоксаль, пиридоксамин)
В основе структуры витамина В6 лежит пиридиновое кольцо. Известны 3 формы витамина В6, отличающиеся строением замещающей группы у атома углерода в n-положении к атому азота. Все они обладают одинаковой биологической активностью:
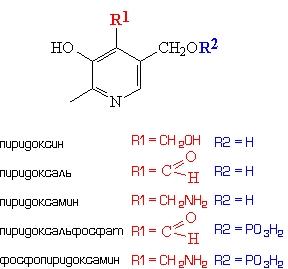

Витамин В6 в виде пиридоксальфосфата и пиридоксаминфосфата, для образования которых расходуется АТР при участии фермента пиридоксалькиназы, выполняет коферментную функцию. Пиридоксалевые ферменты играют ключевую роль в обмене аминокислот, катализируя реакции трансаминирования и декарбоксилирования. Выявлена каталитическая функция пиридаксальфосфата в действии фосфорилазы, играющей, как известно, центральную роль в метаболизме гликогена в организме.
Витамин В6 широко распространен в природе, синтезируется растениями и микроорганизмами, в том числе и микрофлорой кишечника. Однако того количества витамина В6, которое продуцируется микроорганизмами недостаточно для полного обеспечения витамином организма человека. Поэтому основным источником пиридоксина являются продукты питания. Наиболее богаты витамином В6 сухие дрожжи, печень, почки, сердце, мясо, рыба, цельное зерно злаковых и их отруби, горох, бобы, свежий зеленый перец.
Суточная потребность составляет 2-3 мг.
Витамин В6 относится к антидерматитным витаминам. Недостаточность витамина В6 сопровождается дерматитами, стоматитами, глосситами, конъюктивитами, гипохромной анемией, задержкой роста. Авитаминоз В6 у детей проявляется повышенной возбудимостью. Развитие гиповитаминоза этого витамина может быть связано не только с недостаточным поступлением его в организм, но и с нарушением фосфорилирования пиридоксина в желудочнокишечном тракте при заболеваниях органов пишеварения.
Витамин в9 (фолиевая кислота)
Фолиевая кислота состоит из трех структурных единиц: остатка птеридина (I), парааминобензойной кислоты (II) и глутаминовой кислоты (III).

Фолиевая (птероилглутаминовая) кислота
Витамин, полученный из разных источников, может содержать от трех до шести остатков глутаминовой кислоты. Фолиевая кислота метаболически неактивна, но после восстановления птеридинового кольца может превращяться в тетрагидрофолевую кислоту, обладающую коферментными свойствами ферментов, ответственных за перенос одноуглеродных групп (таких, как формил, метил, метилен, оксиметил). Присоединение одноуглеродных остатков к тетрагидрофолиевой кислоте происходит с помощью ковалентной связи. Эти коферменты участвуют в синтезе пуриновых нуклеотидов, в превращении dUMP в dTMP, обмене глицина и серина.
Источниками фолиевой кислоты служат свежие овощи: салат, шпинат, капуста, лук, помидоры, морковь. Из продуктов животного происхождения наиболее богаты фолиевой кислотой печень, почки, яичный желток, сыр, а также пивные и пекарские дрожжи.
Суточная потребность в фолиевой кислоте варьирует от 50 до200 мкг; из-за плохой всасываемости этого витамина рекомендуемая суточная доза – 400 мкг. Наиболее характерными признаками авитаминоза фолиевой кислоты являются нарушение кроветворения, вызывающее малокровие (макроцитарная анемия), наблюдаются нарушения деятельности органов пищеварения, органов размножения и кожи. Этот авитаминоз может возникнуть в случае подавления микрофлоры кишечника лекарственными препаратами сульфаниламидной природы – структурными аналогами парааминобензойной кислоты и/или при нарушении всасывания витамина в желудочно-кишечном тракте при его заболевании.
Тесная связь фолиевых коферментов с метаболизмом нуклеиновых кислот объясняет существенную роль витамина В9 в жизнедеятельности организма. Некоторые производные птеридина тормозят пролиферацию клеток, в том числе и опухолевых, применяются для подавления опухолевого роста у онкологических больных.