

Рибосомальные рнк
Они принимают участие в образовании рибонуклеопротеинов, формирующих немембранные комплексы – рибосомы. Клетки прокариот и эукариот содержат рибосомы, имеюшие общий план строения. В рибосомы входят высокомолекулярные рРНК, дающие начало 30S-40S- и 50S-60S-субчастицам рибосом; рРНК взаимодействуют с мРНК и аминоацил-тРНК в процессе трансляции. 5S-рРНК выступает в роли посредника между пептидилтрансферазным центром и доменом белкового фактора трансляции, обладающим GTP-азной активностью.
Рибосомальные РНК содержат несколько модифицированных нуклеотидов. Чаще всего это метильные производные азотистых оснований или рибозы. Вторичная структура рРНК характеризуется спирализацией самой на себя полирибонуклеотидной цепи. Биспиральные и линейные участки этих молекул формируют постоянные вариабельные домены, которые затем укладываются в более компактные структуры более высокого порядка (рис. 11.7).
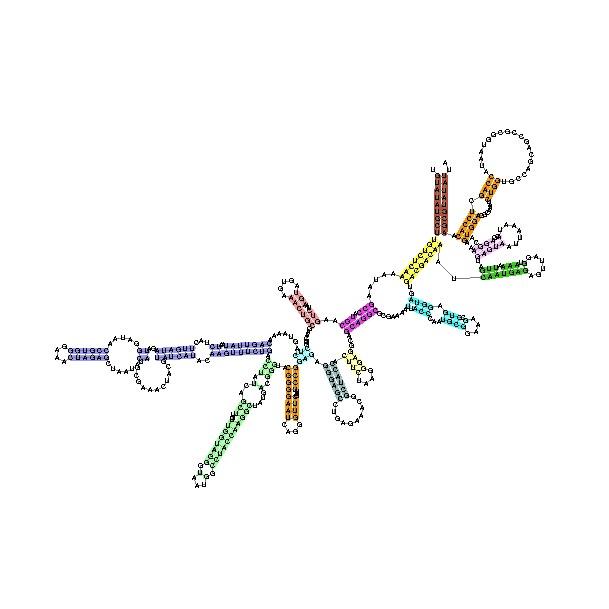
Рис. 11.7. Рибосомальная РНК
Остановимся на малых ядерных РНК. В ядрах всех эукариотических клеток содержится множество коротких и стабильных молекул РНК. Они локализованы в сплайсосомах млекопитающих и называются U-РНК из-за необычайно большого содержания урацила и его модифицированных форм.
Нуклеотидные последовательности всех U-РНК позвоночных совпадают на 95%. В сплайсосому, кроме пяти типов U-РНК, обладающих ферментативной активностью и названных рибозимами, входят еще 50 типов белков. Характерной чертой рибозимов является наличие липких концов, комплементарных концам интронов. Сплайсосома вырезает интроны – участки первичных транскриптов, не несущих информацию о белке.
В последние годы большой интерес исследователей направлен на изучение феномена малых РНК (siRNA). Различают цитоплазматические и ядерные малые РНК. В класс малых (коротких РНК) включают молекулы, содержащие от 20 до 300 нуклеотидов. Самые короткие из них – siRNA состоят из 21 – 28, а млекопитающих из 21 – 23 нуклеотидов. Особенностью этих молекул является то, что они в отличие от большинства других клеточных РНК, состоящих всего из одной цепи нуклеотидов, являются двунитчатыми. Нуклеотиды с противоположных цепей siRNA спариваются друг с другом по тем же законам комплементарности, которые формируют двунитчатые цепи в ДНК. Кроме того, по краям каждой из цепей siRNA всегда остаются два неспаренных нуклеотида.
Ядерным малым РНК отводится способность подавлять экспрессию генов у животныъх и растений. Эффект «гашения» определенных генов малыми РНК получил в молекулярной биологии название РНК-интерференции. Имеются доказательства, что у растений и низших животных организмов siRNA являются элементами «внутриклеточного иммунитета», позволяют быстро распознать и уничтожить чужую РНК.
Лекция 12 витамины – биологическая роль, классификация. Водорастворимые витамины
Витамины представляют собой сборную в химическом отношении группу низкомолекулярных органических веществ, жизненно необходимых для сбалансированного питания. Витамины не синтезируются в организме человека и животных или синтезируются, но в малых количествах, тканями, а также микрофлорой кишечника, присущей организму. Это недостаточно для нормальной жизнедеятельности. Для человека основными источниками витаминов являются высшие растения.
Между витаминами и другими составляющими частями пищи существуют тесные взаимоотношения, объясняемые общностью, единством обмена веществ. В животном мире имеется видовое различие в потребности в отдельных витаминах, что связано с возможностью или невозможностью их достаточного синтеза в организме. Так, аскорбиновая кислота является витамином для человека, обезьяны и морских свинок, тогда как крысы и собаки синтезируют его в процессе промежуточного обмена веществ.
В норме суточная потребность в витаминах мала, однако на потребность в витаминах могут существенно влиять увеличение физической нагрузки, интенсивность умственного труда, физиологическое состояние, возраст, пол, условия окружающей среды.
Поступая в организм с пищей, витамины (большинство из них) выполняют коферментную роль в ферментативных реакциях обмена. Кроме того, они являются компонентами биологическиактивных веществ, выступают в роли антиоксидантов. Анализ структуры коферментов позволяет выделить два функциональных центра, один из которых ответствен за связь с белком, а другой принимает участие непосредственно в каталитическом акте.
В витаминах нуждаются растения, которым эти вещества также необходимы для нормального развития и роста.
В ряде случаев в организм поступают предшественники витаминов, т.н. провитамины, которые в организме превращаются в активные формы витаминов.
Недостаток поступления витаминов с пищей, нарушение всасывания или их использования организмом приводит к развитию патологического состояния – первичные авитаминозы и гиповитаминозы. Напротив, чрезмерное потребление пищевых витаминных форм и/или несбалансированное питание может вызвать гипервитаминозное состояние, которое также является патологическим.
В медицинской и биологической литературе витамины подразделяются на две группы: растворимые в воде и растворимые в жирах. Отдельным витаминам присваивается буквенная, химическая и физиологическая номенклатура.
Жирорастворимые витамины приведены втаблице. 12.1.
Таблица 12.1
Жирорастворимые витамины
-
Буквенное обозначение
Наименование
Физиологическое
действие
А
Ретинол
Антиксерофтальмический
D
Эргокальциферол
Антирахитический
Е
Токоферол
Антистерильный
К
Филлохинон
Антигемморагический
Водорастворимые витамины приведены в таблице 12.2
Таблица 12.2
Водорастворимые витамины
|
Буквенное обозначение |
Наименование |
Физиологическое действие |
|
В1 |
Тиамин |
Антиневритный |
|
В2 |
Рибофлавин |
Витамин роста |
|
В3 (РР) |
Никотиновая кислота, никотинамид
|
Антипеллагрический |
|
В5 |
Пантотеновая кислота |
Антидерматитный |
|
В6 |
Пиридоксин |
Антидерматитный |
|
В9 |
Фолиевая кислота |
Антианемический |
|
В12 |
Цианкобаламин |
Антианемический |
|
Н |
Биотин |
Антисеборейный |
|
С |
Аскорбиновая кислота |
Антискорбутный |
|
Р |
Рутин |
Капилляроукрепляющий |
|
U |
S-метилметионин |
Противоязвенный |
Раскрытие причин авитаминозов и механизма их действий на организм обосновало использование витаминов как лекарственных средств. По лечебно-профилактическому действию была дана следующая групповая характеристика некоторых витаминов. Витамины В1, В2, В3, В5, А и С регулируют функциональное состояние центральной нервной системы, обмен веществ и трофику тканей, поэтому их используют как препараты, повышающие общую реактивность организма. Витамины С, Р, К обеспечивают нормальную проницаемость и устойчивость кровеносных сосудов, повышают свёртываемость крови, т.е. обладают антигеморрагическим эффектом. Витамины В9, В12, С нормализуют и стимулируют кроветворение; их используют как антианемические препараты. Витамины С и А повышают устойчивость организма к инфекциям путем стимулирования синтеза антител и противоспалительных веществ, усиления защиты эпителиев. Витамины А, В2 и С усиливают остроту зрения, расширяют поле цветного зрения.
Среди витаминов есть «отношения» синергизма и антагонизма. Так, влияние витамина Р на проницаемость кровеносных сосудов усиливает витамин С; витамин А снижает токсическое действие антирахитического витамина D, что усиливает эффект последнего. Никотиновая кислота тормозит липотропное действие холина.
В отличие от витаминов есть вещества, обладающие антивитаминными свойствами. Примером может служить тиамин, имеющий высокую структурную специфичность. Если в тиамине изменить радикалы, образуется вещество вытесняющее тиамин из фермента, коферментом которого он является. Антивитаминами являются многие антибиотики и сульфаниамидные препараты.