

Третичная структура белков. Типы нековалентных связей, стабилизирующих третичную структуру. Роль s-s-мостиков в формировании третичной структуры некоторых белков
Под третичной структурой понимают пространственное расположение полипептидной цепи (способ укладки цепи в определенном объеме). В стабилизации пространственной структуры основную роль играют нековалентные связи. К ним относятся водородные связи, электростатические взаимодействия заряженных групп, межмолекулярные ван-дер-ваальсовы силы, взаимодействия неполярных боковых радикалов аминокислот (гидрофобные взаимодействия), диполь-дипольные взаимодействия. Кроме того, важную роль в формировании третичной структуры играют дисульфидные связи (S-S-мостики) (рис.6.4).
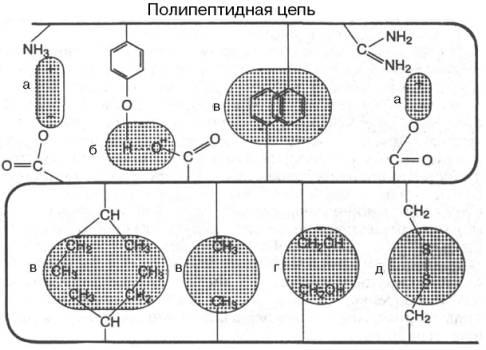
Рис.6.4. Связи, стабилизирующие третичную структуру белка: а) электростатические силы; б) водородная связь; в) гидрофобное взаимодействие; г) диполь-дипольные взаимодействия; д) дисульфидная связь
Дисульфидные связи образуются при окислении сближенных в пространственной структуре белка остатков цистеина в остатки цистина. Считают, что дисульфидные связи, часто множественные, особенно важны для стабилизации маленьких белков, в которых не может возникнуть обширной системы нековалентных взаимодействий.
Третичная структура – уникальное для каждого белка расположение в пространстве полипептидной цепи, зависящее от количества и чередования аминокислот, т.е. предопределенное первичной структурой белка. Конфигурация белковых молекул может быть фибриллярной и глобулярной. Третичная структура многих белков составляется из нескольких компактных глобул, называемых доменами. Между собой домены обычно бывают связаны тонкими перемычками ‒ вытянутыми аморфными полипептидными цепями (рис.6.5).
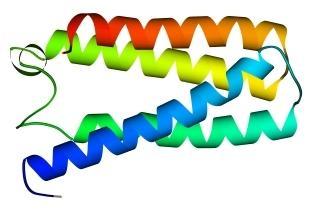


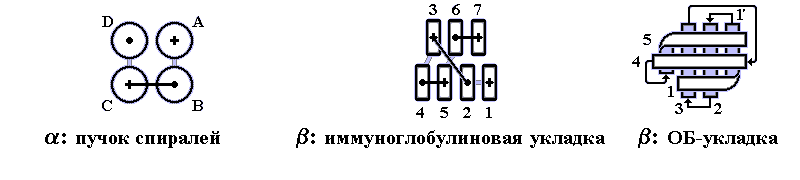
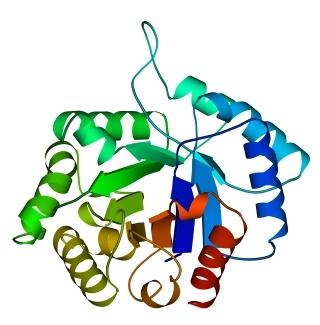
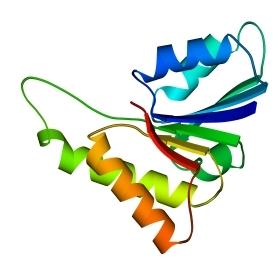
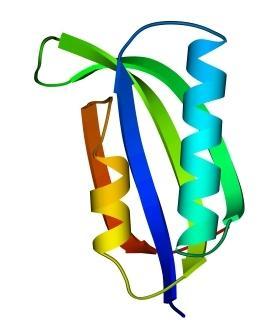
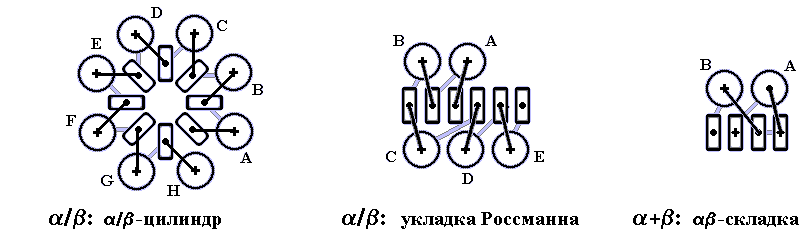
Рис.6.5. Характерные мотивы укладки белковой цепи
Кроме того, в белках встречаются мотивы укладки полипептидной цепи, похожие на орнаменты на индейских и греческих вазах: мотив меандра, мотив греческого ключа, мотив зигзага-"молнии" (рис. 6.6).
При свертывании белковой глобулы значительная часть (не менее половины) гидрофобных радикалов аминокислотных остатков оказывается скрытой от контакта с окружающей белок водой. Происходит образование своеобразных внутримолекулярных «гидрофобных ядер». В них особенно представлены объемные остатки лейцина, изолейцина, фенилаланина, валина.
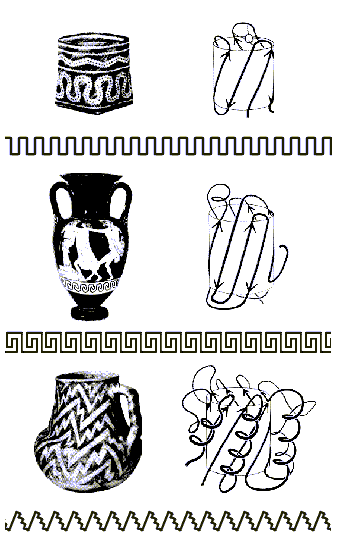
Рис.6.6. Мотивы укладки белковой цепи и орнаменты на индейских и греческих вазах. Вверху: мотив меандра; в середине: мотив греческого ключа; внизу: мотив зигзага-"молнии"
С появлением третичной структуры у белка появляются новые свойства – биологические. В частности, проявление каталитических свойств связано с наличием у белка третичной структуры. И наоборот, нагревание белков, приводящее к разрушению третичной структуры (денатурация), приводит и к утрате биологических свойств.
Четвертичная структура белков. Количество и типы субъединиц.
Заимодействия между субъединицами, стабилизирующие четвертичную структуру. Функциональное значение четвертичной структуры белков
Четвертичная структура − это надмолекулярное образование, состоящее из двух и более полипептидных цепей, связанных между собой нековалентно, а водородными связями, электростатическими, диполь-дипольные и гидрофобными взаимодействиями между остатками аминокислот, находящихся на поверхности. Примером может служить молекула гемоглобина, вирус табачной мозаики (2130 субъединиц).
Каждый из белков-участников третичной структуры при образовании четвертичной структуры называют субъединицей или протомером. Образовавшуюся молекулу называют олигомером, или мультимером. Олигомерные белки чаще построены из четного количества протомеров с одинаковыми или разными молекулярными массами. В образовании четвертичной структуры белка принимают участие те же связи, что и при образовании третичной структуры, за исключением ковалентных.
Объединение белковых молекул третичной структуры без появления новых биологических свойств называют агрегированным состоянием. Как четвертичная структура, так и агрегированное состояние могут быть обратимо разрушены с применением детергентов, в частности, додецилсульфата натрия или неионных детергентов типа тритона. Очень часто для разрушения четвертичной структуры исследуемый белок нагревают при 100С в присутствии 1%-ного 2-меркаптоэтанола и 2%-ного додецилсульфата натрия. В таких условиях восстанавливаются -S-S-связи между остатками Cys, которые в некоторых случаях удерживают субъединицы четвертичной структуры.
Субъединицы, образующие четвертичную структуру белка, могут быть различными как по строению, так и по функциональным свойствам (гетеромеры). Это позволяет объединить в одной структуре несколько взаимосвязанных функций, создать полифункциональную молекулу. Например, в протеинкиназе, стехиометрия червертичной структуры которой отвечает формуле С2R2, субъединица С ответственна за ферментативную активность, осуществляя перенос фосфатного остатка от АТР на белок; субъединица R является регуляторной. В отсутствие циклического АМР последняя связана с С-субъединицей и ингибирует ее. При образовании комплекса с сАМР четвертичная структура распадается и С-субъединицы оказываются способными фосфорилировать белковые субстраты. В гомомерных белках субъединицы одинаковы.
Подавляющая часть белков, имеющих четвертичную структуру, приходится на димеры, тетрамеры и гексамеры, последние встречаются у белков с молекулярной массой, большей 100 кДа.
Характерной особенностью белков с четвертичной структурой является их способность к самосборке. Взаимодействие протомеров осуществляется с высокой специфичностью, благодаря образованию десятка слабых связей между контактными поверхностями субъединиц, поэтому ошибки при формировании четвертичной структуры белков исключены.
Практически все белки-ферменты имеют четвертичную структуру и состоят, как правило, из четного числа протомеров (двух, четырех, шести, восьми). Четвертичная структура белка подразумевает такое объединение белков третичной структуры, при котором появляются новые биологические свойства, не характерные для белка в третичной структуре. В частности, такие эффекты, как кооперативный и аллостерический, характерны лишь для белков с четвертичной структурой.
Четвертичная структура – последний уровень в организации белковой молекулы, причем не обязательный – до половины известных белков ее не имеют.