

Вторичная структура белков: -спираль, ее основные характеристики, -структура, -изгиб. Роль водородных связей в формировании вторичной структуры. Сверхвторичные (надвторичные) структуры белка
Вторичная структура − это пространственное расположение полипептидной цепочки в виде α-спирали или β-складчатости безотносительно к типам боковых радикалов и их конформации.
Л. Полинг и Р. Кори предложили модель вторичной структуры белка в виде α-спирали, в которой водородные связи замыкаются между каждой первой и четвертой аминокислотой, что позволяет сохранять нативную структуру белка, осуществлять простейшие функции, защищать от разрушения. В образовании водородных связей принимают участие все пептидные группы, что обеспечивает максимальную стабильность, снижает гидрофильность и увеличивает гидрофобность белковой молекулы. α-спираль образуется самопроизвольно и является наиболее устойчивой конформацией, отвечающей минимуму свободной энергии.
Наиболее распространенным элементом вторичной структуры является правая α-спираль (αR). Пептидная цепь здесь изгибается винтообразно. Ha каждый виток приходится 3,6 аминокислотного остатка, шаг винта, т.е. минимальное расстояние между двумя эквивалентными точками, составляет 0,54 нм; α-спираль стабилизирована почти линейными водородными связями между NH-группой и СО-группой четвертого по счету аминокислотного остатка. Таким образом, в протяженных спиральных участках каждый аминокислотный остаток принимает участие в формировании двух водородных связей. Неполярные или амфифильные α-спирали с 5-6 витками часто обеспечивают заякоривание белков в биологических мембранах (трансмембранные спирали). Зеркально-симметричная относительно αR-спирали левая α-спираль (αL) встречается в природе крайне редко, хотя энергетически возможна. Закручивание полипептидной цепи белка в спиралеобразную структуру происходит вследствие взаимодействия между кислородом карбонильной группы i-того аминокислотного остатка и водородом амидогруппы (i+4)- аминокислотного остатка посредством образования водородных связей (рис.6.1).

Рис. 6.1. Вторичная структура белка: α-спираль
Другая форма спирали присутствует в коллагене, важнейшем компоненте соединительных тканей. Это левая спираль коллагена с шагом 0,96 нм и при остатке в 3,3 в каждом витке более пологая по сравнению с α-спиралью. В отличие от α-спирали образование водородных мостиков здесь невозможно. Структура стабилизирована за счет скручивания трех пептидных цепей в правую тройную спираль.
Наряду с α-спиралями в образовании вторичной структуры белка принимают также участие β-структуры, β-изгиб.
В отличие от конденсированной α-спирали β-слои почти полностью вытянуты и могут располагаться как параллельно, так и антипараллельно (рис.6.2).
![]()
![]()
а) б)
Рис.6.2. Параллельное (а) и антипараллельное (б) расположение β-слоев
B складчатых структурах также образуются поперечные межцепочечные водородные связи (рис.6.3). Если цепи ориентированы в противоположных направлениях, структура называется антипараллельным складчатым листом (βα); если цепи ориентированы в одном направлении, структура называется параллельным складчатым листом (βn). В складчатых структурах α-С-атомы располагаются на перегибах, а боковые цепи ориентированы почти перпендикулярно средней плоскости листа, попеременно вверх и вниз. Энергетически предпочтительной оказывается βα-складчатая структура с почти линейными H-мостиками. В растянутых складчатых листах отдельные цепи чаще всего не параллельны, а несколько изогнуты относительно друг друга.
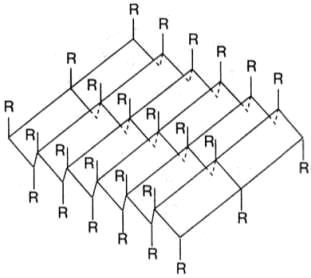
Рис.6.3. β-складчатая структура
Кроме регулярных в полипептидных цепях есть еще и нерегулярные вторичные структуры, т.е. стандартные структуры, не образующие длинных периодических систем. Это – β-изгибы они называются так потому, что часто стягивают верхушки соседних β-тяжей в антипараллельных β-шпильках). В изгибы обычно входит около половины остатков, не опавших в регулярные структуры белков.
Супервторичная структура − это более высокий уровень организации белковой молекулы, представленный ансамблем взаимодействующих между собой вторичных структур:
1. α-спираль − два антипараллельных участка, которые взаимодействуют гидрофобными комплементарными поверхностями (по принципу «впадина-выступ»);
2. сверхспирализация α-спирали;
3. βхβ − два параллельных участка β-цепи;
4. β-зигзаг.
Встречаются разнообразные способы укладки белковой цепи (рис. 6.5). Рисунок 6.5 взят с обложки журнала Nature 1977 г. (v.268, №.5620), где была напечатана статья Дж. Ричардсона о мотивах укладки белковых цепей.
Домен – компактная глобулярная структурная единица внутри полипептидной цепи. Домены могут выполнять разные функции и подвергаться свертыванию в независимые компактные глобулярные структурные единицы, соединенные между собой гибкими участками внутри белковой молекулы.