

Номенклатура пептидов и полипептидов. Природные пептиды: глутатион, карнозин, ансерин, грамицидин s, окситоцин, энкефалины
Пептиды ‒ природные или синтетические соединения, молекулы которых построены из остатков α-аминокислот, соединенных пептидными (амидными) связями. Пептиды могут содержать также неаминокислотную компоненту. По числу аминокислотных остатков, входящих в молекулы пептидов, различают дипептиды, трипептиды, тетрапептиды и т.д. Пептиды, содержащие до десяти аминокислотных остатков, называются олигопептидами, содержащие более десяти аминокислотных остатков – полипептидами. Природные полипептиды с молекулярной массой более 6000 называются белками.
Аминокислотный остаток пептидов, несущий свободную α-аминогруппу, называется N-концевым, а остаток, несущий свободную α-карбоксильную группу – С-концевым. Название пептида образуется из названий входящих в его состав аминокислотных остатков, перечисляемых последовательно, начиная с N-концевого. При этом используют тривиальные названия аминокислот, в которых суффикс "ин" заменяется на "ил". Исключение составляет C-концевой остаток, название которого совпадает с названием соответствующей аминокислоты. Все аминокислотные остатки, входящие в пептиды, нумеруются, начиная с N-конца. Для записи первичной структуры пептида (аминокислотной последовательности) широко используют трех- и однобуквенные обозначения аминокислотных остатков (например, Ala-Ser-Asp-Phe-GIy – это аланил-серил-аспарагил-фенилаланил-глицин).
Отдельные представители пептидов
Глутатион — трипептид -глутамилцистеинилглицин, содержащийся во всех животных и растительных клетках, бактериях.
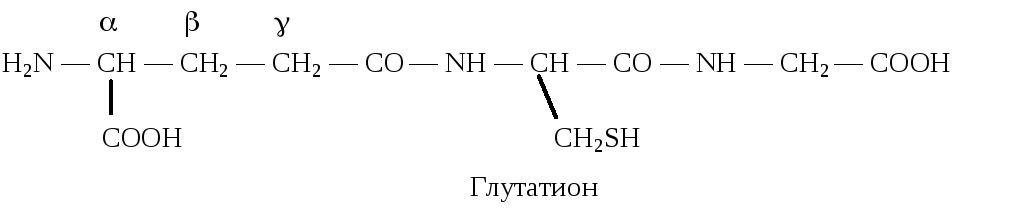
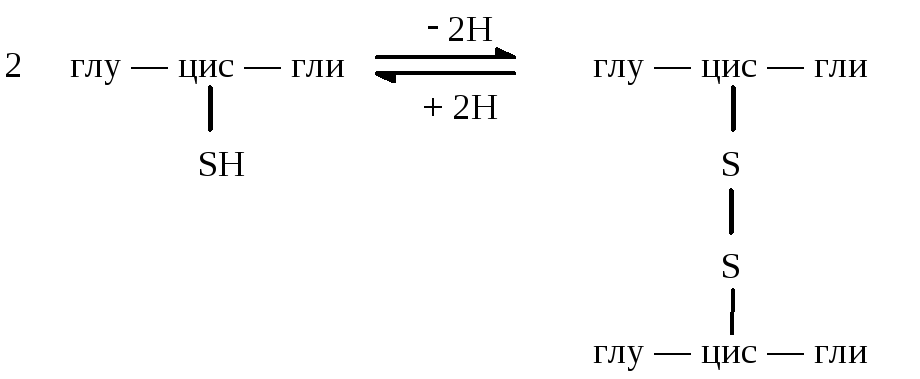
Глутатион участвует в ряде окислительно-восстановительных процессов. Он выполняет функцию антиоксиданта. Это обусловлено наличием в его составе цистеина и определяет возможность существования глутатиона в восстановленной и окисленной формах.
Карнозин (от лат. carnosus ‒ мясной, caro ‒ мясо), C9H14O3N4, – дипептид (β-аланилгистидин), состоящий из аминокислот β-аланина и L-гистидина. Открыт в 1900 г. В. С. Гулевичем в мясном экстракте. Молекулярная масса 226, кристаллизуется в виде бесцветных игл, хорошо растворим в воде, нерастворим в спирте. Содержится в скелетной мускулатуре большинства позвоночных. Среди рыб встречаются виды, у которых карнозин и составляющие его аминокислоты отсутствуют (либо присутствует только L-гистидин или только β-аланин). В мышцах беспозвоночных карнозина нет. Содержание карнозина в мышцах позвоночных колеблется обычно от 200 до 400 мг% их сырой массы и зависит от их структуры и функции; у человека ‒ около 100-150 мг%.
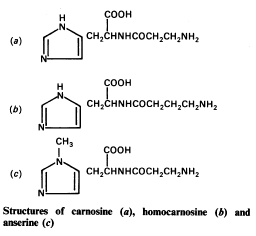
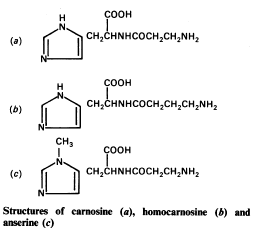
Карнозин (β-аланил-L-гистидин) Ансерин (β-аланил-1-метил- L-гистидин)
Влияние карнозина на биохимические процессы, протекающие в скелетных мышцах, разнообразно, однако окончательно биологическая роль карнозина не установлена. Добавление карнозина к раствору, омывающему мышцу изолированного нервно-мышечного препарата, вызывает восстановление сокращений утомлённой мышцы.
Дипептид ансерин (N-метилкарнозин или β-аланил-1-метил- L-гистидин), сходный по строению с карнозином, в мышцах человека отсутствует, но имеется в скелетных мышцах тех видов, мышцы которых способны к быстрым сокращениям (мышцы конечностей кролика, грудная мышца птиц). Физиологические функции β-аланил-имидазольных дипептидов не вполне ясны. Возможно, они выполняют буферные функции и поддерживают рН в скелетной мышце, сокращающейся в анаэробных условиях. Однако ясно, что карнозин и ансерин стимулируют АТР-азную активность миозина in vitro, увеличивают амплитуду мышечного сокращения, предварительно сниженную утомлением. Академик С.Е. Северин показал, что имидазолсодержащие дипептиды не влияют непосредственно на сократительный аппарат, но увеличивают эффективность работы ионных насосов мышечной клетки. Оба дипептида образуют хелатные комплексы с медью и способствуют поглощению этого металла.
Антибиотик грамицидин S выделен из Bacillus brevis и представляющий собой циклический декапептид:
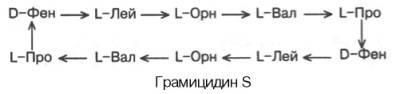
Грамицидин S
В структуре грамицидина S имеются 2 остатка орнитина, производные аминокислоты аргинина и 2 остатка D-изомеров фенилаланина.
Окситоцин − гормон, вырабатываемый нейросекреторными клетками передних ядер гипоталамуса и затем переносимый по нервным волокнам в заднюю долю гипофиза, где он накопляется и откуда выделяется в кровь. Окситоцин вызывает сокращение гладких мышц матки и в меньшей степени − мышц мочевого пузыря и кишечника, стимулирует отделение молока молочными железами. По химической природе окситоцин − октапептид, в молекуле которого 4 остатка аминокислот связаны в кольцо цистином, соединённым также с трипептидом: Pro-Leu-Gly.
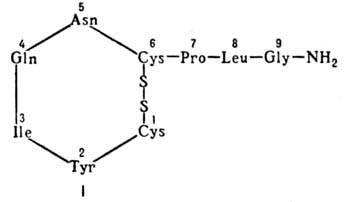
окситоцин
Рассмотрим нейропептиды (опиатные пептиды). Первые два представителя нейропептидов, названные энкефалинами, были выделены из мозга животных:
Тир — Гли — Гли — Фен — Мет- Мет-энкефалин
Тир — Гли — Гли — Фен — Лей- Лей-энкефалин
Эти пептиды обладают обезболивающим действием и используются как лекарственные средства.