

Зовнішня регуляція активності транскрипційних факторів
Активність певного гена залежить від наявності у клітині певного набору активаторів / репресорів транскрипції. Відповідно, гени самих факторів транскрипції перебувають під контролем складних систем регуляції, що працюють під час розвитку та диференціації клітин.
У результаті в клітині певного типу відбувається синтез специфічного набору ТФ, що призводить до активації специфічного набору генів. У той же час, експресія певного гена може оперативно контролюватися у відповідь на зовнішні сигнали шляхом зміни активності вже синтезованих транскрипційних факторів.
Два найважливіші механізми такої регуляції – взаємодія ТФ певного типу зі стероїдними гормонами та каскади посттрансляційних модифікацій у відповідь на дію хімічних сигналів (сигнальна трансдукція).
Гормонові рецептори – транскрипційні фактори, активність яких залежить від стероїдних гормонів. Гормоновий рецептор (зазвичай гомодимер) складається з трьох структурних доменів: 1) ДНК-зв’язувальний домен, що має специфічну спорідненість до певних елементів послідовності ДНК; 2) активаційний домен; 3) гормон-зв’язувальний домен. У відсутності гормону білок міститься в цитоплазмі, де гормон-зв’язувальний домен взаємодіє з білком теплового шоку hsp90, який підтримує недоструктурований стан домену.
У результаті гормоновий рецептор є інактивованим (рис. 13).
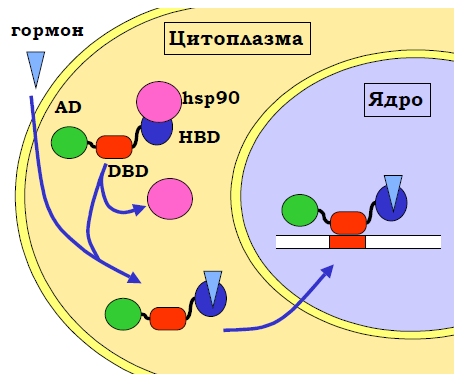
Рис. 13. Активація гормонового рецептора стероїдним гормоном. AD – активаційний, DBD – ДНК-зв’язувальний, HBD – гормон-зв’язувальний домени.
Коли гормон проникає в цитоплазму, він взаємодіє з гормон-зв’язувальним доменом, витісняючи hsp90, відбувається остаточне структурування гормонового рецептора, і той стає активним – прямує до ядра, де зв’язується зі специфічним елементом послідовності. Сайт зв’язування гормонового рецептора є досить маленьким, а чим коротшою є довжина сайта зв’язування, тим імовірнішою є його доступність. Тому взаємодія гормонового рецептора з ДНК часто запускає каскад збирання енхансосоми: рецептор рекрутує інші транскрипційні фактори за рахунок свого активаційного домену.
Сигнальна трансдукція. Приклад сигнальної трансдукції наведено на рис. 14. Білковий гормон не проникає у клітину, а зв’язується рецептором на зовнішньому боці мембрани. Рецептор є гомодимером, і у відсутності гормону дві субодиниці вільно дифундують у площині мембрани. Зв’язування гормону викликає димеризацію, унаслідок чого в цитоплазматичної частини рецептора виникає кіназна активність. Кіназа фосфорилює неактивні субодиниці транскрипційного фактора.
Після фосфорилювання субодиниці об’єднуються в димер, що й активує цей фактор: він проникає в ядро, де знаходить специфічну послідовність ДНК. Наведена схема є найпростішим варіантом сигнальної трансдукції: часто примембранна кіназа запускає каскад фосфорилювання – фосфорилює білок, який набуває внаслідок цього кіназної активності, ця нова кіназа фосфорилює інший білок (або кілька різних білків, завдяки чому здійснюється підсилення сигналу та / або його розгалуження по кількох шляхах, спрямованих до кількох кінцевих мішеней), перетворюючи його на кіназу, і так далі – до фосфорилювання та, відповідно, активації транскрипційного фактора.

Рис. 14. Активація транскрипційного факторау відповідь на дію білкового гормону.