

Ініціація трансляції в еукаріотів
Еукаріотична рибосома поза процесом трансляції також дисоційована на дві субодиниці, завдяки взаємодії маленької з фактором IF3 (складається з 9 – 11 субодиниць). Крім того, дисоційований стан субодиниць підтримується кількома іншими факторами, що взаємодіють з маленькою (eIF3С) і великою (eIF6) субодиницями. З комплексом маленька субодиниця – eIF3 зв’язується потрійнийкомплекс ініціаторна метіонінова тРНК – eIF2·GTP (eIF2 є аналогом фактора елонгації eEF1).
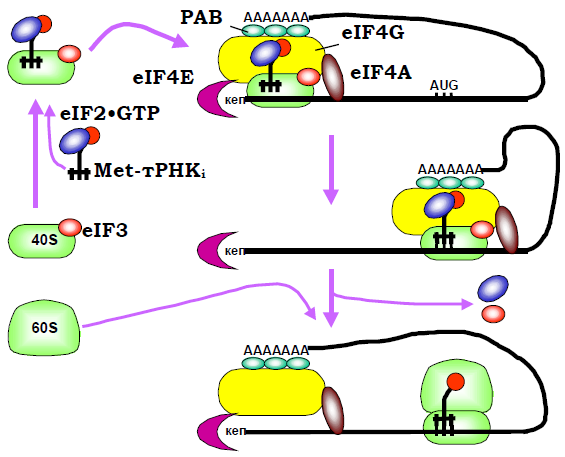
Рис. 28. Послідовність основних подій при ініціації трансляції в еукаріотів.
Паралельно (рис. 28) відбувається впізнання кепа на 5′-кінці мРНК факторами ініціації групи eIF4: субодиниця eIF4E зв’язуєтьсяз кепом, eIF4G є структурним модулем, що забезпечує цілісність комплексу та його взаємодію з іншими елементами ініціації, eIF4Aє АТР-залежною РНК-геліказою, фактор eIF4В (не показаний на рис. 28) взаємодіє з мРНК, стимулює АТР-азну активність гелікази і при скануванні мРНК бере участь у впізнанні стартового кодона.
Наступним кроком є об’єднання кеп-асоційованого комплексу з таким, що асоційований з маленькою субодиницею (ініціаторна тРНК має бути присутньою на маленькій субодиниці, щоб ця остання могла приєднатися до кепу). Ефективність утворення цього преініціаторного комплексу на кепі підсилюється за рахунок взаємодії polyA-зв’язаних білків (PABP) з eIF4G: polyA-послідовність на 3′-кінці мРНК також приєднується до преініціаторного комплексу, замикаючи мРНК у кільце. Далі за рахунок геліказної активності eIF4A преініціаторний комплекс (залишаючи на кепі лише eIF4E) починає транслокацію: eIF4A здійснює гідроліз АТР і пересувається вздовж мРНК у напрямку до 3′-кінця, одночасно руйнуючи дволанцюгові шпильки, якщо вони зустрічаються. Під час транслокації преініціаторний комплекс сканує послідовність мРНК, перевіряючи її на наявність стартового кодона.
Зовсім не обов’язково при цьому перший кодон AUG, що зустрічається, сприймається як стартовий. Упізнання стартового кодона залежить від контексту послідовності, в якій він розташований. Найкращим контекстом, який максимально сприяє ініціації трансляції, є так звана послідовність Козак (Marilyn Kozak): GCC(A/G)CCAUGG. Упізнання стартового кодона викликає відповідні конфірмаційні зміни комплексу, звільнення факторів eIF3 і eIF4 (останній повертається на кеп разом polyA-ділянкою мРНК), приєднання великої рибосомної субодиниці та дисоціацію eIF2·GDP після гідролізу GTP.
Дисоціація eIF3 і eIF2 полегшується фактором eIF5. Рибосома після цього вступає в етап елонгації трансляції, а на кепі починається збирання наступної рибосоми. Таким чином, на мРНК одночасно працює кілька рибосом, які утворюють так звану полісому (рис. 29). Зациклення полісоми: 1) сприяє залученню зруйнованої при термінації трансляції у 3′-кінцевій зоні мРНК рибосоми до нового раунду ініціації; 2) забезпечує додатковий захист 3′-кінця мРНК (унаслідок його взаємодії з кеп-асоційованим комплексом) від нуклеазної деградації; 3) підвищує надійність системи трансляції – оскільки деградація мРНК може розпочинатися тільки з 3′-кінця, лише повноцінні матриці, що містять polyA-послідовність (і автоматично – кодуючу частину) залучаються до білкового синтезу.
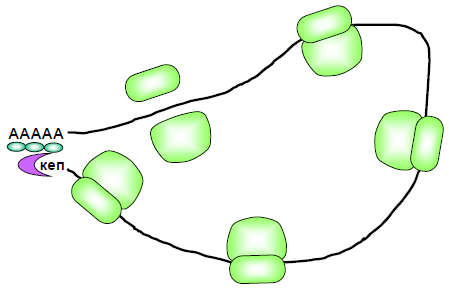
Рис. 29. Циркулярна полісома.