

Лимитирующие стадии.
Как уже обсуждалось в выше, традиционный кинетический подход для описания скорости химических реакций в микродисперсных системах (мицеллах, микроэмульсиях и т.д.) имеет ряд существенных ограничений. Обсуждению одного из них, связанного со стохастическим характером процессов в малых системах, была посвящена лекция 3. Другое важное ограничение состоит в том, что скорость химической реакции в таких системах, как правило, лимитируется скоростью процессов межмицеллярного обмена, а не скоростью самой реакции в объеме микрореактора (т.е. в объеме самой мицеллы). Следовательно, в результате реакции функция распределения молекул реагентов по мицеллам будет существенно отличаться от простого случайного распределения. В этом случае центр тяжести кинетического описания химического процесса переносится на рассмотрение особенностей механизма межмицеллярного обмена.
Сделаем оценку характерных времен химической реакции, протекающей внутри мицеллы и сравним их с соответствующими величинами, определяющими скорости процессов межмицеллярного обмена. Можно считать, что скорость реакции определяется диффузией реагирующих молекул. В предыдущей лекции вероятность элементарного акта химической реакции характеризовалась величиной k, физический смысл которой есть средняя частота встречи двух частиц в единицу времени. Она зависит как от характеристик самой мицеллы так и от свойств реагирующих частиц. Сделаем оценку величины k для мицеллы. Рассмотрим две молекулы, диффундирующие внутри мицеллы, представляющей собой сферу радиуса Rm, и пусть r0 есть сумма радиусов этих молекул. Предположим, что одна молекула зафиксирована в центре мицеллы, другая диффундирует по ее объему. Характерное значение времени встречи двух молекул объеме мицеллы, находящихся в начальный момент времени на расстоянии r друг от друга, можно оценить как среднее время достижения внутренней границы при диффузионном движении точечной частицы, в пространстве ограниченном двумя коаксиальными сферами, где радиус внешней сферы равен радиусу мицеллы, а радиус внутренней сферы равен сумме радиусов взаимодействующих молекул.
Дифференциальное уравнение для нахождения имеет вид
![]() , (1)
, (1)
при следующих граничных условиях:
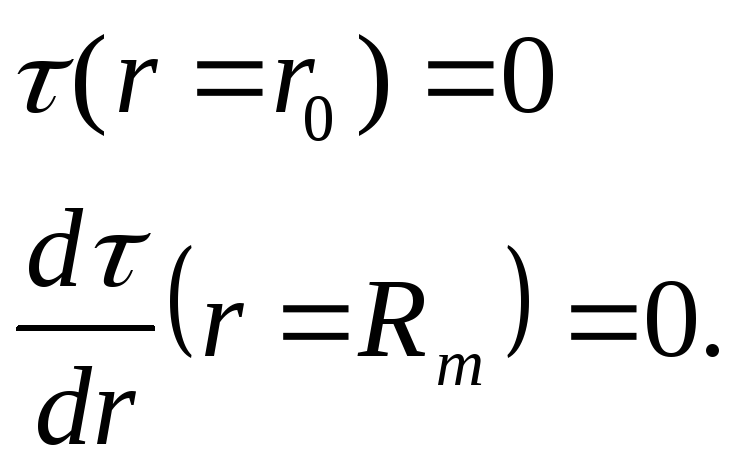 (2)
(2)
где D – коэффициент взаимной диффузии реагирующих молекул. Второе граничное условие соответствует отражающей стенке, которой является граница мицеллы.
Решением уравнения (1), при граничных условиях (2) будет
![]() (3)
(3)
Усредняя
по всем возможным начальным значениям
r,
получим,
что при условии
![]()

![]() (4)
(4)
Для характерных значений параметров реальных мицеллярных систем и микроэмульсий, которые могут изменяться в довольно широких пределах (D~10-510-7 см2/сек, Rm ~110 нм), получаем, что величина k может изменяться в пределах некольких порядков 105 108 сек-1.
Для сравнения сделаем оценку характерных времен межмицеллярного обмена реагирующих молекул. Предположим, что обмен происходит в результате столкновений мицелл друг с другом. Можно оценить частоту соударений мицелл в растворах как произведение характерного значения концентрации мицелл на константу скорости диффузионно-контролируемого процесса их столкновений. Число мицелл в единице объема может изменяться в широких пределах в зависимости от концентрации ПАВ. Можно в качестве приближенного среднего значения взять отношение ККМ к числу агрегации мицеллы. Характерные значения для концентрации мицелл имеет порядок 10-310-5 моль/л, и, умножая эту величину на характерное значение константы скорости диффузионно-контролируемых процессов в растворах, т.е. на величину порядка 1091010 моль-1л сек-1, получаем значение частоты межмицеллярного обмена в пределах 103107сек-1. Обмен солюбилизированными молекулами может также происходить непосредственно через дисперсионную среду. В этом случае характерные частоты столкновений молекул, находящихся в среде с отдельно взятой мицеллой также определяется произведением константы скорости диффузионно-контролируемого процесса на концентрацию молекул в среде. Учитывая, что, как правило, солюбилизированные молекулы являются лиофобными по отношению к дисперсионной среде, т.е. их концентрация в среде не превышает значения концентрации мицелл, для частоты обмена через среду получаются те же значения, что и при межмицеллярном обмене.