

Электрические и химические синапсы
Электрические и химические синапсы [c.188] Передача действия нервного импульса на другие клетки происходит в специальных образованиях, которые называются синапсами. Синапсы — это места контакта нервного окончания с поверхностной мембраной регулируемой клетки. Они включают пресинаптическую и постсинаптическую мембраны, атакже синаптическую щель (рис. 110). Выделяют электрические и химические синапсы. В химических синапсах передача нервного импульса происходит с участием нейромедиаторов (нейротрансмиттеров). Нейромедиаторами являютсятакие химические вещества, как ацетил-холин, адреналин, норадреналин, а также дофамин, серотонин, глутамин, глицин, ГАМК и др. Синапс приспособлен к быстрому выбросу нейромедиатора, образующегося в эфферентных нервных клетках в синаптических пузырьках. Поэтому нервная система оказывает быстрое воздействие на довольно ограниченный участок органа. Поскольку скелетные мышцы регулируются химическими синапсами с участием нейропередатчика ацетил- [c.277] Первым таким признаком является временная задержка, те самые 0,8 мс в случае электрического синапса ждать задержки не было оснований. Другим отличительным свойством химических реакций является их сильная температурная зависимость. Чем выше температура (в определенных пределах), тем быстрее идет реакция. В случае злектрических синапсов ожидать такой зависимости также не приходилось. Кроме того, при изучении химическихсинапсов было показано, что для их работы нужны ионы кальция в среде, омывающей клетки. Замена кальция на магний блокировала передачу сигналачерез химический синапс. И вот в 1957 г. с помощью микроэлектродов был Открыт синапс, в котором сигнал передавался практически без задержки, передача слабо зависела от температуры п не блокировалась магнием. Был открыт первыйчисто электрический синапс. Это показалось странным исключением, да и синапс открыли всего лишь у какого-то речного рака. Но лиха беда начало. Вскореэлектрические синапсы открыли у рыб KomeKj обезьян... [c.158] Самый простой способ передачи сигнала от нейрона к нейрон - это прямое электрическое сопряжение через щелевые контакты. Главное преимуществотаких электрических синапсов состоит в том. что сигналы передаются без задержки. С другой стороны, эти синапсы гораздо меньще приспособлены для регулирования и адаптации, чем химические синапсы, с помощью которых осуществляется большинство связей между нейронами. Электрическая связь через щелевые контакты была рассмотрена в гл. 14 (разд. 14.1.5-14.1.8), здесь же речь пойдет только о химических синапсах. [c.304] ТО, как мы видели, убедительными свидетельствами того, что существуют какхимические синапсы, так и электрические (а также смешанные). [c.205] Передача сигналов от клетки к клетке. может осуществляться либо путем прямого прохождения потенциалов действия (электрические синапсы), либо с помощью специальных молекул — нейромедиаторов (химические синапсы). В зависимости от своих специфических функций синапсы имеют очень разные структуры. Вхимических синапсах расстояние между клетками составляет - 20—40 нмсинаптическая щель между клетками— это часть межклеточного пространстваона содержит жидкость с низким электрическим сопротивлением, так чтоэлектрический сигнал рассеивается прежде, чем он достигнет следующей клетки.Электрическая передача, напротив, осуществляется только в специализированных структурах — щелевых контактах, где клетки находятся на расстоянии 2 нм и соединяются проводящими канала.ми. В действительности здесь имеется нечто сходное с постулированным ранее синцитием, или многоклеточным цитоплазматическим континуумом. По иронии истории нау- [c.188] Трансмиттер-зависимые ионные каналы приспособлены для превращения внеклеточных химических сигналов в электрические сигналы. Они располагаются обычно в специализированных соединениях (называемых химическими синапсами), расположенных между нервными клетками и клетками-мишенями. Эти каналы концентрируются на плазматической мембране клетки-мишени в области синапса. Каналы способны открываться на некоторое время в ответ на связывание нейротрансмиттера, высвобождаемого нервным окончанием. При этом меняется проницаемость постсинаптической мембраны клетки-мишени(рис. 6-62). В отличие от потенциал-зависимых каналов, ответственных за возникновение потенциалов действия, трансмиттер-зависимые каналы относительно нечувствительны к мембранном) потенциалу и поэтому неспособны к самоусиливающемуся возбуждению. Вместо этого они изменяют проницаемость мембраны и, следовательно, влияют на мембранный потенциал. Величина этогоизменения зависит от того, сколько трансмиттера высвободилось в синапсе и в течение какого времени он там присутствует. Ясно, что потенциал действия можетвозникнуть только при условии, что потенциал-зависимые каналы также присутствуют в этой же мембране клетки-мишени. [c.402] Возможно и сочетание обоих механизмов, электрического и химического, в одном смешанном синапсе, однако в нервной системе млекопитающих преобладаютчисто химические синапсы. [c.207] В отличие от электрических синапсов, которые так или иначе служат дляобразования непрерывного пути для электрического сигнала, химические синапсы — это настоящие разрывы электрическому сигналу напрямую через химический синапс не пройти. [c.163] Аналогичная ситуация имеется и с химическими синапсами. Принцип ихработы используется в организмах не только для передачи информации, но и вдругих целях. Оказывается, разнообразные секреторные клетки используют ионы Са + для регуляции выброса секрета подобно тому, как в химическом синапсе этот процесс используется для выброса медиатора. Кроме того, клетки многих желез являются электрически возбудимыми. [c.176] Ответ на этот вопрос был получен в работах исследователей из Стэнфордского университета Джона Николлса и его сотрудников. Рассмотрим сначала Ь-нейрон. В тело этого нейрона и в тело сенсорного нейрона, расположенного в том же ганглии, были введены микроэлектроды. Прямое раздражение сенсорного лейрона путемподачи электрического импульса через внутриклеточный электрод приводило к возникновению в этом нейроне потенциала действия. Это сопровождалось и реакцией со сто-,роны Ь-нейрона, которая заключалась в деполяризующем синаптическом потенциале, вызывавшем возникновение небольшого поамплитуде импульса (см. рис. 20.2). Анализ латентных периодов и других свойств потенциалов, возникавших при стимуляции всех трех разновидностейсенсорных нейронов, показал, что Т-нейрон соединен с Ь-нейроном электрическим синапсом, Ы-нейрон — химическим синапсом, а Р-нейрон — и электриче- Ским, и химическим синапсами. Было обнаружено также, чтохимические синапсы обладают высокой пластичностью при повторном раздражении ответ в них существенно облегчается. Напротив, ответы, опосредуемыеэлектрическими синапсами, оставались сравнительно неизменными (рис. 20.2). [c.52] Химический синапс (163). Выделение медиатора (164). Работапостсинаптической мембраны (167). Какие синапсы лучше — электрические или химические (169) Химический синапс и торможение (171). О величине синаптических потенциалов (173). Устройства, подобные синапсам (174). Нервная клетка — клетка (177) [c.285] В некоторых тканях роль такого сопряжения очевидна. Например,электрическое сопряжение синхронизирует сокращения клеток сердечной мышцы и клеток гладкой мускулатуры, ответственных за перистальтику кишечника. Аналогичным образом электрические синапсы между нейронамипозволяют потенциалам действия быстро распространяться с клетки на клетку без задержки, происходящей в химических синапсах это дает преимущество в случаях, когда решающее значение имеют быстрота и надежность реакции, например принекоторых реакциях бегства у рыб и насекомых. [c.217] Нервные сигналы переходят от клетки к клетке через синапсы, которые могут быть электрическими (щелевые контакты) или химическими. В химическом синапсе деполяризация пресинаптической мембраны в результате прибытиянервного импульса открывает потенциал-зависимые кальциевые каналы, вызывая тем самым приток Са в клетку, что приводит к освобождению нейромедиатора из синаптических пузырьков. Медиатор диффундирует всинаптическую щель и связывается с рецепторными белками в мембранепостсинаптической клетки в конечном итоге медиатор удаляется изсинаптической щели путем диффузии, ферментативного расщепления или обратного поглощения выделившей его клеткой. Через рецепторные белки,образующие лиганд-зависимые каналы, реализуется быстрый постсинаптический эффект нейромедиатора-открытие каналов приводит к возникновению возбуждающего или тормозного постсинаптического потенциам в соответствии сионной специфичностью каналов. При участии рецепторов, сопряженных с ферментог ми, например с аденилатциклазой, обычно осуществляются медленные и более продолжительные эффекты. [c.111] Скелетные мышечные волокна позвоночных, подобно нервным клеткам, способны возбуждаться под действием электрического тока, и поэтому нервно-мышечное соединение (рис. 19-16) оказалось хорошей моделью химического синапса вообще. Двигательный нерв и иннервируемую им мышц> можно отделить от окружающей ткани и поддерживать в функционально активном состоянии впитательной среде определенного состава. Стимулируя нерв через наружные электроды, можно с помощью внутриклеточного микроэлектрода регистрировать ответ одиночной мышечной клетки (рис. 19-17). Па рис. 19-18 сравниваетсятонкая структура нервно-мышечного соединения и типичного синапса межд> двумя нейронами центральной нервной системы. [c.305] Специальные преобразователи превращают сенсорные стимулы в электрические сигналы. Например, у позвоночных волосковые клетки внутреннего уха представляют собой механорецепторы на свободной поверхности каждойволосковой клетки имеется пучок стереоцилий (гигантских микроворсинок), и при наклоне таких пучков открываются ионные каналы, что ведет к изменениюмембранного потенциала Мембранный потенциал фоторецепторных клеток вглазу позвоночного изменяется при поглощении света молекулами родопсина, содержащимися в этих клетках. И в том, и в другом случае электрический сигнал, возникающий в сенсорной клетке вначале в форме рецепторного потенциала, передается соседним нейронам через химические синапсы. Однако два упомянутых класса клеток-рецепторов используют для выработки рецепторных потенциалов различные стратегии в основе одной лежат рецепторные молекулы, связанные с каналами, а другая зависит от молекул-рецепторов, не связанных с каналами. Вволосковых клетках физическая связь между стереоцилиями порождаетмеханические силы, которые прямо воздействуют на ионные каналы в плазматической мембране, заставляя их быстро открываться или закрываться. Впалочках сетчатки активированные светом молекулы родопсина инициируюткаскад ферментативных реакций, в результате которых в цитозоле гидролизуется цикличе- [c.345]
|
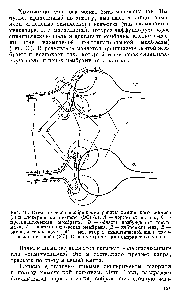
Рис. 40. Схематическое изображение работы химического синапса (а) и электрического синапса(ЭС) (6). А — терминаль аксона, Б —пресинаптическая мембрана, В — область возбуждения терлв-нали, Г — постсинаптическая мембрана, Д — нейрон-мишень, Е — синаптическая щель, М медиатор в синаптической щелихимического синапса (ХС). Стрелками показано направление токов |
|
 <=""
img="" style="border: none; display: block;
margin-left: auto; margin-right: auto;">
<=""
img="" style="border: none; display: block;
margin-left: auto; margin-right: auto;">Вклад ионов кальция в создание потенциалов действия может иметь важное значение. Во-первых, вход кальция во время потенциала действия представляетсобой эффективный механизм повышения внутриклеточной концентрациисвободного Са +, а этот ион участвует в работе целого ряда клеточных механизмов. Во-вторых, ионы кальция регулируют проницаемость для других ионов, в частности для К+. В-третьих, Са+ играет важнейшую роль в модуляции проведения в электрических синапсах и в выделении медиаторов вхимических синапсах. Подробнее мы рассмотрим эти механизмы как впоследующих разделах настоящей главы, так и в главе 8. [c.162] Электрические синапсы сравнительно редки, и их роль в центральной нервной системе высших организмов пока неясна. Фершпан и Поттер открыли их в брюшном нерве краба, а позднее их обнаружили в многочисленных организмах моллюсках,, членистоногих и млекопитающих. В противоположность химическому синапсу, где прохождение импульса несколько задерживается из-за высвобождения и диффузии медиатора, сигнал через электрический синапспередается быстро. Физиологическая важность таких синапсов может, следовательно, быть связана с необходимостью быстрого сопряжения специфических клеток. [c.189] Самый простой способ передачи сигнала от нейрона к нейрону-это прямое электрическое взаимодействие через щелевые контакты. Такие электрпескк сшишсы между нейронами встречаются в некоторых участках нервной системы у многих животных, в том числе и у позвоночных. Главное преимущество электрических синапсов состоит в том, что сигнал передается без задержки. С другой стороны, эти синапсы не приспособлены к выполнению некоторых функций и не могут так тонко регулироваться, как химические синапсы, через которые осуществляется большинство связей между нейронами. Электрическая связьчерез щелевые контакты была рассмотрена в главе [c.93] Скелетные мышечные волокна позвоночных, подобно нервным клеткам, способны возбуждаться под действием электрического тока, и нервно-мышечноесоедшенае (рис. 18-24) может служить хорошей моделью химического синапсавообще. На рис. 18-25 сравнивается тонкая структура этого синапса с типичным синапсом между двумя нейронами головного мозга. Двигательный нерв и иннервируемую им мышцу можно отделить от окрузкаюшей ткани и поддерживать вфункционирующем состоянии в среде определенного состава. Возбуждая нерв через наружные электроды, можно с помошью внутриклеточного микроэлектрода регистрировать ответ одиночной мышечной клетки (рис. 18-26). Микроэлектрод сравнительно легко ввести в волокно скелетной мышцы, так как это очень крупная клетка (порядка 100 мкм в диаметре). [c.96]
|
Рис. 42. Строение химического синапса иэквивалентная схема, поясняющая механизм его действия а — Слева терминаль аксона (Л) ипресинаптическая мембрана с кальциевыми каналами (Г) внутри синаптического утолщения видны везикулы, наполненные медиатором (В), и митохондрии (Б). Е — синаптическая щель, разделяющая синаптическое утолщение и клетку-мишонь. В ност-синаптическую мембрану (Д) встроены молекулы белка-рецептора, каналы которого открываются придействии медиатора, б — Эквивалентная электрическая схема клетки-мишени С — емкость мембраны, й общ — проводимость несинаптической мембраны, V — источник э.д.с., создающий ПП, — проводимость постсинаптической мембраны, которая сильно растет при действии медиатора |
|
 <=""
img="" style="border: none; display: block;
margin-left: auto; margin-right: auto;">
<=""
img="" style="border: none; display: block;
margin-left: auto; margin-right: auto;">В стволе мозга млекопитающих были найдены три области с электрическими синапсами. Все они приведены на рис. 8.3. В мезенцефальном ядре тройничного нерва электрические синапсы образуются между телами соседних клеток, атакже между телами и начальными сегментами аксонов (А). В вестибулярном ядре Дейтерса такие синапсы образуются между телами клеток и окончаниями аксона.Потенциал действия, возникающий в одной из клеток, через короткийлатентный период вызывает деполяризацию в соседней клетке ток, приводящий к этой деполяризации, проходит через окончания аксона (рис. 8.3Б). В нижнейоливе электрические синапсы образуются между шипиками дендритов. На этих шипиках имеются и химические синапсы полагают, что активация последних приводит к шунтированию электрических синапсов и разобщению клеток (этот гипотетичный механизм представлен на рис. 8.3В и Г). [c.178] Различные типы клеточных контактов перечислены в табл. 12-1. Основной тип адгезионных контактов назьтают десмосомами. Плотные контакты (и септированные контакты у беспозвоночных)-главные виды замыкающих контактов. Проводящие контакты бьтают двух видов щелевые контакты и химические синапсы. Через щелевой контакт малые молекулы могут непосредственно переходить из одной клетки в другую, а в химическом синапсе клетки не имеютнепосредственной связи, хотя и очень сближены (рис. 12-18). Входная клетка синапса (пресинаптическая) вьщеляет вещество (нейромедиатор), котороедиффундирует через синаптическую щель и вызывает реакцию другой, постсинаптической, клетки. Так как химические синапсы подробно рассмотрены в главе 18, мы не будем останавливаться на них здесь. Химические синапсы не следует смешивать с более редкими электрическими синапсами, в которыхэлектрические импульсы непосредственно переходят с одной нервной клеткина другую через щелевой контакт. [c.211] Нервные клетки, или нейроны, - это клетки с необычайно длинными отростками, передающими электрические сигналы в виде потенциалов Оействия - бегущих волн электрического возбужОения. Обычно от тела нервной клеткиотхоОит несколько разветвленных дендритов и один длинный аксон. Как правило, сигналы воспринимаются дендритами и телом клетки, а затем распространяются по аксону и передаются другим клеткам в химических синапсах. Здесьэлектрический сигнал, приходящий в пресинаптическое окончание аксона, индуцирует секрецию нейромедиатора, который в свою очередь вызываетэлектрическое изменение в постсинаптической клетке [c.294] Нервные сигналы передаются от клетки к клетке через синапсы, которые могут быть электрическими (щелевые контакты) или химическими. В химическом синапсе в результате деполяризации пресипаптической мембраны поо действием нервного импульса открываются потенциал-зависимые кальциевые каналы, что приводит к притоку ионов Са, которые в свою очередь вызываютвысвобождение нейромедиатора из синаптических пузырьков путем экзоцитоза. Медиатор диффундирует через синаптическую щель и связываетсярецепторными белками в мембране постсинаптической клетки. Изсинаптической щели медиатор быстро удаляется путем диффузии,ферментативного расщепления или ж всасывания окончанием аксона илиглиальными клетками. Рецептори, пейромедиаторов можно подразделить на связанные и не связанные [c.318] В гл.7 уже упоминалось разделение синапсзав на химические, электрические и смешанные. Чем вьппе степень эволюционной организации нервной системы, тем разнообразиее природа химических синапсов. Особенно это касается головного мозга высших млекопитающих, включая человека. Очевидно, химические синапсыоказались эволюционно более выгодными для передачи дискретных сигналов по сравнению с другими типами межклеточных контактов, поскольку на их основе возможна не только передача сигнала, но и его разнообразная модуляция, в том числе гуморальными факторами. Основой восприятия нейроном химического сигнала в синапсе, а также ряда модулирующих влияний являются рецепторы. [c.255] В некоторьк тканях роль сопряжения клеток через щелевые контактыочевидна. Например, электрическое сопряжение синхронизирует сокращения клеток сердечной мышцы и клеток гладкой мускулатуры, ответственных заперистальтику кишечника. Точно так же электрическое сопряжение между нервными клетками позволяет потенциалам действия быстро распространяться от клетки к клетке без задержки, происходящей в химических синапсах это дает преимущество в случаях, когда решающее значение имеют быстрота и надежность ответа, например при некоторых реакциях бегства у рыб и насекомых. Труднее понять, зачем нужны щелевые контакты в тканях, не проявляюшцхэлектрической активности. В принципе обмен метаболитами и ионами мог бы обеспечить координацию активности отдельных клеток в этих тканях. Например, через щелевые контакты могла бы координироваться такая активность клетокэпителиального слоя, как биение ресничек а поскольку внутриклеточные посредники типа циклического АМР способны проходить через щелевые контакты, ответ сопряженных клеток на внеклеточные сигнальные молекулы мог бы распространяться и координироваться именно этим пугем. [c.484] Мембраны многих клеток способны возбуждаться под действием специфических химических или физических стимулов. Ответы мембраны аюгонанервной клетки на электрический стимул, синапса на выделение медиатора,палочек сетчатки на свет, подвижных клеток на молекулы аттрактан-тов-все это реакции, опосредованные возбудимыми комплексами, присутствующими в мембранах. Эти и другие процессы, опосредованные возбудимыми комплею ами, имеютследующие общие особенности. [c.326] Нейроны характеризуются необыкновенно высоким уровнем обмена веществ, значительная часть которого направлена на обеспечение работы натриевого насоса в мембранах и поддержание состояния возбуждения. Химические основыпередачи нервного импульса по аксону уже обсуждались в гл. 5, разд. Б, 3. Последовательное раскрытие сначала натриевых и затем калиевых каналов можно считать твердо установленным. Менее ясным остается вопрос, сопряжено лиизменение ионной проницаемости, необходимое для распространения потенциала действия, с какими-либо особыми ферментативными процессами. Нахманзон указывает, что ацетилхолинэстераза присутствует в высокой концентрации на всем протяжении мембраны нейрона, а не только в синапсах [38, 39]. Он предполагает, что увеличение проницаемости к ионам натрияобусловлено кооперативным связыванием нескольких молекул ацетилхолина смембранными рецепторами, которые либо сами составляют натриевые каналы, либо регулируют степень их открытия. При этом ацетилхолин высвобождается из участков накопления, расположенных на мембране, в результате деполяризации. Собственно, последовательность событий должна быть такова, что изменение электрического поля в мембране индуцирует изменение конформации белков, а это уже приводит к высвобождению ацетилхолина. Под действием аце-тилхолинэстеразы последний быстро распадается, и проницаемость мембраныдля ионов натрия возвращается к исходному уровню. В целом приведенное описание отличается от описанной ранее схемы синаптической передачи только в одном отношении в нейронах ацетилхолин накапливается в связанной с белками форме, тогда как в синапсах — в специальных пузырьках. Существует мнение, что работа калиевых каналов регулируется ионами кальция. Чувствительный к изменению электрического поля Са-связывающий белок высвобождает Са +, который в свою очередь активирует каналы для К" , последнее происходит с некоторым запозданием относительно времени открытия натриевых каналов, что обусловлено различием вконстантах скоростей этих двух процессов [123]. Закрытие калиевых каналов обеспечивается энергией гидролиза АТР. Имеются и другие предположения омеханизмах нервной проводимости [124]. Некоторые из них исходят из того, что нервная проводимость целиком обеспечивается работой натриевого насоса. [c.349]