

Торможение (физиология)
Торможение — в физиологии — активный нервный процесс, вызываемый возбуждением и проявляющийся в угнетении или предупреждении другой волны возбуждения. Обеспечивает (вместе с возбуждением) нормальную деятельность всех органов и организма в целом. Имеет охранительное значение (в первую очередь для нервных клеток коры головного мозга), защищая нервную систему от перевозбуждения.
И. П. Павлов называл иррадиацию торможения по коре больших полушарий головного мозга «проклятым вопросом физиологии».
Центральное торможение
Центральное торможение открыто в 1862 г. И. М. Сеченовым. В процессе опыта он удалил у лягушки головной мозг на уровне зрительных бугров и определял время сгибательного рефлекса. Затем на зрительные бугры помещался кристалл соли в результате чего наблюдалось увеличение продолжительности времени рефлекса. Это наблюдение позволило И. М. Сеченову высказать мнение о явлении торможения в ЦНС. Данный тип торможения называют сеченовским или центральным.
Ухтомский объяснил результаты с позиции доминанты. В зрительных буграх — доминанта возбуждения, которая подавляет действие спинного мозга.
Введенский объяснил результаты с позиции отрицательной индукции. Если в центральной нервной системе возникает возбуждение в определенном нервном центре, то вокруг очага возбуждения индуцируется торможение. Современное объяснение: при раздражении зрительных бугров возбуждается каудальный отдел ретикулярной формации. Эти нейроны возбуждают тормозные клетки спинного мозга (клетки Реншоу), которые тормозят активность альфа-мотонейронов спинного мозга.
Первичное торможение
Первичное торможение возникает в специальных тормозных клетках, примыкающих к тормозному нейрону. При этом тормозные нейроны выделяют соответствующие нейромедиаторы.
Виды первичного торможения
Постсинаптическое — основной вид первичного торможения, вызывается возбуждением клеток Реншоу и вставочных нейронов. При этом типе торможения происходит гиперполяризация постсинаптической мембраны, что и обуславливает торможение. Примеры первичного торможения:
Возвратное — нейрон воздействует на клетку, которая в ответ тормозит этот же нейрон.
Реципрокное — это взаимное торможение, при котором возбуждение одной группы нервных клеток обеспечивает торможение других клеток через вставочный нейрон.
Латеральное — тормозная клетка тормозит расположенные рядом нейроны. Подобные явления развиваются между биполярными и ганглиозными клеткамисетчатки, что создает условия для более четкого видения предмета.
Возвратное облегчение — нейтрализация торможения нейрона при торможении тормозных клеток другими тормозными клетками.
Пресинаптическое — возникает в обычных нейронах, связано с процессом возбуждения.
Вторичное торможение
Вторичное торможение возникает в тех же нейронах, которые генерируют возбуждение.
Виды вторичного торможения
Пессимальное торможение — это вторичное торможение, которое развивается в возбуждающих синапсах в результате сильной деполяризации постсинаптической мембраны под действием множественной импульсации.
Торможение вслед за возбуждением возникает в обычных нейронах и также связано с процессом возбуждения. В конце акта возбуждения нейрона в нем может развиваться сильная следовая гиперполяризация. В то же время возбуждающий постсинаптический потенциал не может довести деполяризацию мембраны докритического уровня деполяризации, потенциалзависимые натриевые каналы не открываются и потенциал действия не возникает.
Периферическое торможение
Открыто братьями Вебер в 1845 г. В качестве примера можно привести торможение деятельности сердца (снижение ЧСС) при раздражении блуждающего нерва.
Условное и безусловное торможение
Термины «условное» и «безусловное» торможение предложены И. П. Павловым.
Условное торможение
Условное, или внутреннее, торможение — форма торможения условного рефлекса, возникающее при неподкреплении условных раздражителей безусловными. Условное торможение является приобретенным свойством и вырабатывается в процессе онтогенеза. Условное торможение является центральным торможением и ослабевает с возрастом.[1]
Безусловное торможение
Безусловное (внешнее) торможение — торможение условного рефлекса, возникающее под действием безусловных рефлексов (например, ориентировочного рефлекса). И. П. Павлов относил безусловное торможение к врожденным свойствам нервной системы, то есть безусловное торможение является формой центрального торможения.[1]
Торможение
Координирующая функция локальных нейронных сетей помимо усиления может выражаться и в ослаблении слишком интенсивной активности нейронов за счет их торможения.
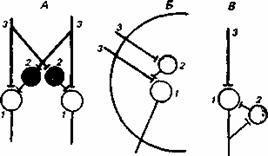
Рис.8.1 .Реципрокное (А), пресинаптическое (Б) и возвратное (В) торможение в локальных нейронных цепях спинного мозга
1 - мотонейрон; 2 - тормозный интернейрон; 3 - афферентные терминали.
Торможение, как особый нервный процесс, характеризуется отсутствием способности к активному распространению по нервной клетке и может быть представлено двумя формами - первичным и вторичным торможением.
Первичное торможение обусловлено наличием специфических тормозных структур и развивается первично без предварительного возбуждения. Примером первичного торможения является так называемое реципрокное торможение мыщц-антагонистов, обнаруженное в спинальных рефлекторых дугах . Суть этого явления состоит в том, что если активируются проприорецепторы мышцы-сгибателя, то они через первичные афференты одновременно возбуждают мотонейрон данной мышцы-сгибателя и через коллатераль афферентного волокна тормозный вставочный нейрон. Возбуждение вставочного нейрона приводит к постсинаптическому торможению мотонейрона антагонистической мышцы-разгибателя, на теле которого аксон тормозного интернейрона формирует специализированные тормозные синапсы. Реципрокное торможение играет важную роль в автоматической координации двигательных актов.
Другим примером первичного торможения является открытое Б. Реншоу возвратное торможение. Оно осуществляется в нейронной цепи, которая состоит из мотонейрона и вставочного тормозного нейрона - клетки Реншоу. Импульсы от возбужденного мотонейрона через отходящие от его аксона возвратные кол-латерали активируют клетку Реншоу, которая в свою очередь вызывает торможение разрядов данного мотонейрона. Это торможение реализуется за счет функции тормозных синапсов, которые клетка Реншоу образует на теле активирующего ее мотонейрона. Таким образом, из двух нейронов формируется контур с отрицательной обратной связью, позволяющий стабилизировать частоту разрядов моторной клетки и подавить идущую к мышцам избыточную импульсацию.
В ряде случаев клетки Реншоу формируют тормозные синапсы не только на активирующих их мотонейронах, но и на соседних мотонейронах со сходными функциями. Осуществляемое через эту систему торможение окружающих клеток называется латеральным.
Торможение по принципу отрицательной обратной связи встречается не только на выходе, но и на входе моторных центров спинного мозга. Явление подобного рода описано в моносинаптических соединенях афферентных волокон со спинальными мотонейронами, торможение которых при данной ситуации не связано с изменениями в постсинаптической мембране. Последнее обстоятельство позволило определить данную форму торможения как пресинаптическое. Оно обусловлено наличием вставочных тормозных нейронов, к которым подходят коллатерали афферентных волокон . В свою очередь, вставочные нейроны формируют аксо-аксональные синапсы на афферентных терминалях, являющихся пресинаптическими по отношению к мотонейронам. В случае избыточного притока сенсорной информации с периферии происходит активация тормозных интернейронов, которые через аксо-аксональные синапсы вызывают деполяризацию афферентных термина-леи и, таким образом, уменьшают количество выделяемого из них медиатора, а следовательно, и эффективность синаптической передачи. Электрофизиологическим показателем этого процесса является снижение амплитуды регистрируемых от мотонейрона ВПСП. Вместе с тем никаких признаков изменений ионной проницаемости или генерации ТПСП в мотонейронах не наблюдается.
Вопрос о механизмах пресинаптического торможения является довольно сложным. По-видимому, медиатором в тормозном аксо-аксональном синапсе является гамма-аминомасляная кислота, которая вызывает деполяризацию афферентных терминалей за счет увеличения проницаемости их мембраны для ионов С1-. Деполяризация снижает амплитуду потенциалов действия в афферентных волокнах и тем самым уменьшает квантовый выброс медиатора в синапсе. Другой возможной причиной деполяризации терминалей может быть повышение наружной концентрации ионов К+ при длительной активации афферентных входов. Следует отметить, что феномен пресинаптического торможения обнаружен не только в спинном мозгу, но и в других отделах ЦНС.
Исследуя координирующую роль торможения в локальных нейронных цепях, следует упомянуть еще об одной форме торможения - вторичном торможении, которое возникает без участия специализированных тормозных структур как следствие избыточной активации возбуждающих входов нейрона. В специальной литературе эту форму торможения определяют как торможение Введенского, который открыл его в 1886 г. при исследовании нервно-мышечного синапса.
Торможение Введенского играет предохранительную роль и возникает при чрезмерной активации центральных нейронов в полисинаптических рефлекторных дугах. Оно выражается в стойкой деполяризации клеточной мембраны, превышающей критический уровень и вызывающей инактивацию Na-каналов, ответственных за генерацию потенциалов действия. Таким образом, процессы торможения в локальных нейронных сетях уменьшают избыточную активность и участвуют в поддержании оптимальных режимов импульсной активности нервных клеток.
ТОРМОЖЕНИЕ В ЦНС. ВИДЫ И ЗНАЧЕНИЕ.
Проявление и осуществление рефлекса возможно только при ограничении распространения возбуждения с одних нервных центров на другие. Это достигается взаимодействием возбуждения с другим нервным процессом, противоположным по эффекту процессом торможения.
Почти до середины XIX века физиологи изучали и знали только один нервный процесс - возбуждение.
Явления торможения в нервных центрах, т.е. в центральной нервной системе были впервые открыты в 1862 году И.М.Сеченовым ("сеченовское торможение”). Это открытие сыграло в физиологии не меньшую роль, чем сама формулировка понятия рефлекса, так как торможение обязательно участвует во всех без исключения нервных актах. И.М.Сеченов обнаружил явление центрального торможения при раздражении промежуточного мозга теплокровных. В 1880 году немецкий физиолог Ф.Гольц установил торможение спинальных рефлексов. Н.Е. Введенский в результате серий опытов по парабиозу вскрыл интимную связь процессов возбуждения и торможения и доказал, что природа этих процессов едина.
Торможение -местный нервный процесс, приводящий к угнетению или предупреждению возбуждения. Торможение является активным нервным процессом, результатом которого служит ограничение или задержка возбуждения. Одна из характерных черт тормозного процесса- отсутствие способности к активному распространению по нервным структурам.
В настоящее время в центральной нервной системе выделяют два вида торможения:торможение центральное (первичное),являющееся результатом возбуждения (активации) специальных тормозных нейронов иторможение вторичное, которое осуществляется без участия специальных тормозных структур в тех самых нейронах в которых происходит возбуждение.
Центральное торможение(первичное) - нервный процесс, возникающий в ЦНС и приводящий к ослаблению или предотвращению возбуждения. Согласно современным представлениям центральное торможение связано с действием тормозных нейронов или синапсов, продуцирующих тормозные медиаторы (глицин, гаммааминомасляную кислоту), которые вызывают на постсинаптической мембране особый тип электрических изменений, названных тормозными постсинаптическими потенциалами (ТПСП) или деполяризацию пресинаптического нервного окончания, с которым контактирует другое нервное окончание аксона. Поэтому выделяют центральное (первичное) постсинаптическое торможение и центральное (первичное) пресинаптическое торможение.
Постсинаптическое торможение(лат. post позади, после чего-либо + греч. sinapsis соприкосновение, соединение) - нервный процесс, обусловленный действием на постсинаптическую мембрану специфических тормозных медиаторов (глицин, гаммааминомаслянная кислота), выделяемых специализированными пресинаптическими нервными окончаниями. Медиатор, выделяемый ими, изменяет свойства постсинаптической мембраны, что вызывает подавление способности клетки генерировать возбуждение. При этом происходит кратковременное повышение проницаемости постсинаптической мембраны к ионам К+ или CI, вызывающее снижение ее входного электрического сопротивления и генерацию тормозного постсинаптического потенциала (ТПСП). Возникновение ТПСП в ответ на афферентное раздражение обязательно связано с включением в тормозной процесс дополнительного звена - тормозного интернейрона, аксональные окончания которого выделяют тормозной медиатор. Специфика тормозных постсинаптических эффектов впервые была изучена на мотонейронах млекопитающих (Д. Экклс, 1951). В дальнейшем первичные ТПСП были зарегистрированы в промежуточных нейронах спинного и продолговатого мозга, в нейронах ретикулярной формации, коры больших полушарий, мозжечка и таламических ядер теплокровных животных.
Известно, что при возбуждении центра сгибателей одной из конечностей центр ее разгибателей тормозится и наоборот. Д. Экклс выяснил механизм этого явления в следующем опыте. Он раздражал афферентный нерв, вызывающий возбуждение мотонейрона, иннервирующего мышцу - разгибатель.
Нервные импульсы, дойдя до афферентного нейрона в спинномозговом ганглии, направляются по его аксону в спинном мозге по двум путям: к мотонейрону, иннервирующему мышцу - разгибатель, возбуждая ее и по коллатерам к промежуточному тормозному нейрону, аксон которого контактирует с мотонейроном иннервирующим мышцу - сгибатель, вызывая таким образом торможение антагонистической мышцы. Этот вид торможения был обнаружении в промежуточных нейронах всех уровней центральной нервной системы при взаимодействии антагонистических центров. Он был назван поступательным постсинаптическим торможением. Этот вид торможения координирует, распределяет процессы возбуждения и торможения между нервными центрами.
Возвратное (антидромное) постсинаптическое торможение(греч. antidromeo бежать в противоположном направлении) - процесс регуляции нервными клетками интенсивности поступающих к ним сигналов по принципу отрицательной обратной связи. Он заключается в том, что коллатерали аксонов нервной клетки устанавливают синаптические контакты со специальными вставочными нейронами (клетки Реншоу), роль которых заключается в воздействии на нейроны, конвергирующие на клетке, посылающей эти аксонные коллатерали (рис. 87). По такому принципу осуществляетсяторможение мотонейронов.
Возникновение импульса в мотонейроне млекопитающих не только активирует мышечные волокна, но через коллатерали аксона активирует тормозные клетки Реншоу. Последние устанавливают синаптические связи с мотонейронами. Поэтому усиление импульсации мотонейрона ведет к большей активации клеток Реншоу, вызывающей усиление торможения мотонейронов и уменьшение частоты их импульсации. Термин "антидромное” употребляется потому, что тормозной эффект легко вызывается антидромными импульсами, рефлекторно возникающими в мотонейронах.
Чем сильнее возбужден мотонейрон, чем больше сильные импульсы идут к скелетным мышцам по его аксону, тем интенсивнее возбуждается клетка Реншоу, которая подавляет активность мотонейрона. Следовательно, в нервной системе существует механизм, оберегающий нейроны от чрезмерного возбуждения. Характерная особенность постсинаптического торможения заключается в том, что оно подавляется стрихнином и столбнячным токсином (на процессы возбуждения эти фармакологические вещества не действуют).
В результате подавления постсинаптического торможения нарушается регуляция возбуждения в цнс, возбуждение разливается ("диффундирует”) по всей цнс, вызывая перевозбуждение мотонейронов и судорожные сокращения групп мышц (судороги).
Торможение ретикулярное(лат. reticularis - сетчатый) - нервный процесс развивающийся в спинальных нейронах под влиянием нисходящей импульсации из ретикулярной формации (гигантское ретикулярное ядро продолговатого мозга). Эффекты, создаваемые ретикулярными влияниями, по функциональному действию сходны с возвратным торможением, развивающимся на мотонейронах. Влияние ретикулярной формации вызывают стойкие ТПСП, охватывающие все мотонейроны независимо от их функциональной принадлежности. В этом случае, так же как и при возвратном торможении мотонейронов происходит ограничение их активности. Между таким нисходящим контролем со стороны ретикулярной формации и системочй возвратного торможения через клетки Реншоу существует определенное взаимодействие, и клетки Реншоу находятся под постоянным тормозящем контролем со стороны двух структур. Тормозящее влияние со стороны ретикулярной формации являются дополнительным фактором в регуляции уровня активности мотонейронов.
Первичное торможение может вызываться механизмами иной природы, не связанными с изменениями свойств постсинаптической мембраны. Торможение в этом случае возникает на пресинаптической мембране (синаптическое и пресинаптическое торможение).
Синаптическое торможение(греч. sunapsis соприкосновение, соединение) - нервный процесс, основанный на взаимодействии медиатора, секретируемого и выделяемого пресинаптическими нервными окончаниями, со специфическими молекулами постсинаптической мембраны. Возбуждающий или тормозной характер действия медиатора зависит от природы каналов, которые открываются в постсинаптической мембране. Прямое доказательство наличия в цнс специфических тормозящих синапсов было впервые получено Д. Ллойдом (1941).
Данные относительно электрофизиологических проявлений синаптического торможения: наличие синаптической задержки, отсутствие электрического поля в области синаптических окончаний дали основание считать его следствием химического действия особого тормозящего медиатора, выделяемого синаптическими окончаниями. Д. Ллойд показал, что если клетка находится в состоянии деполяризации, то тормозной медиатор вызывает гиперполяризацию, в то время как на фоне гиперполяризации постсинаптической мембраны он вызывает ее деполяризацию.
Пресинаптическое торможение (лат. praе -впереди чего-либо + греч. sunapsis соприкосновение, соединение) - частный случай синаптических тормозных процессов, проявляющихся в подавлении активности нейрона в результате уменьшения эффективности действия возбуждающих синапсов еще на пресинаптическом звене путем угнетения процесса высвобождения медиатора возбуждающими нервными окончаниями. В этом случае свойства постсинаптической мембраны не подвергаются каким-либо изменениям. Пресинаптическое торможение осуществляется посредством специальных тормозных интернейронов. Его структурной основой являются аксо-аксональные синапсы, образованные терминалиями аксонов тормозных интернейронов и аксональными окончаниями возбуждающих нейронов.
При этом окончание аксона тормозного нейрона является пресимпатическим по отношению к терминали возбуждающего нейрона, которая оказывается постсинаптической по отношению к тормозному окончанию и пресинаптической по отношению к активируемой им нервной клетки. В окончаниях пресинаптического тормозного аксона освобождается медиатор, который вызывает деполяризацию возбуждающих окончаний за счет увеличения проницаемости их мембраны для CI. Деполяризация вызывает уменьшение амплитуды потенциала действия, приходящего в возбуждающее окончание аксона. В результате происходит угнетение процесса высвобождения медиатора возбуждающими нервными окончаниями и снижение амплитуды возбуждающего постсинаптического потенциала.
Характерной особенностью пресинаптической деполяризации является замедленное развитие и большая длительность (несколько сотен миллисекунд), даже после одиночного афферентного импульса.
Пресинаптическое торможение существенно отличается от постсинаптического и в фармакологическом отношении. Стрихнин и столбнячный токсин не влияют на его течение. Однако наркотизирующие вещества (хлоралоза, нембутал) значительно усиливают и удлиняют пресинаптическое торможение. Этот вид торможения обнаружен в различных отделах цнс. Наиболее часто оно выявляется в структурах мозгового ствола и спинного мозга. В первых исследованиях механизмов пресинаптического торможения считалось, что тормозное действие осуществляется в точке, отдаленной от сомы нейрона, поэтому его называли "отдаленным” торможением.
Функциональное значение пресинаптического торможения, охватывающего пресинаптические терминали, по которым поступают афферентные импульсы, заключается в ограничении поступления к нервным центрам афферентной импульсации. Пресинаптическое торможение в первую очередь блокирует слабые асинхронные афферентные сигналы и пропускает более сильные, следовательно, оно служит механизмом выделения, вычленения более интенсивных афферентных импульсов из общего потока. Это имеет огромное приспособительное значение для организма, так как из всех афферентных сигналов, идущих к нервным центрам, выделяются самые главные, самые необходимые для данного конкретного времени. Благодаря этому нервные центры, нервная система в целом освобождается от переработки менее существенной информации.
Вторичное торможение- торможение осуществляющееся теми же нервными структурами, в которых происходит возбуждение. Этот нервный процесс подробно изложен в работах Н.Е. Введенского (1886, 1901г.г.).
Торможение реципрокное (лат. reciprocus - взаимный) - нервный процесс, основанный на том, что одни и те же афферентные пути, через которые осуществляется возбуждение одной группы нервных клеток, обеспечивают через посредство вставочных нейронов торможение других групп клеток. Реципрокные отношения возбуждения и торможения в цнс были открыты и продемонстрированы Н.Е. Введенским: раздражение кожи на задней лапке у лягушки вызывает ее сгибание и торможение сгибания или разгибания на противоположной стороне. Взаимодействие возбуждения и торможения является общим свойством всей нервной системы и обнаруживается как в головном, так и в спинном мозге. Экспериментально доказано, что нормальное выполнение каждого естественного двигательного акта основано на взаимодействии возбуждения и торможения на одних и тех же нейронах цнс.
Общее центральное торможение -нервный процесс, развивающийся при любой рефлекторной деятельности и захватывавающий почти всю цнс, включая центры головного мозга. Общее центральное торможение обычно проявляется раньше возникновения какой-либо двигательной реакции. Оно может проявляться при такой малой силе раздражения при которой двигательный эффект отсутствует. Такого вида торможение было впервые описано И.С. Беритовым (1937). Оно обеспечивает концентрацию возбуждения других рефлекторных или поведенческих актов, которые могли бы возникнуть под влиянием раздражений. Важная роль в создании общего центрального торможения принадлежит желатинозной субстанции спинного мозга.
При электрическом раздражении желатинозной субстанции у спинального препарата кошки происходит общее торможение рефлекторных реакций, вызываемых раздражением сенсорных нервов. Общее торможение является важным фактором в создании целостной поведенческой деятельности животных, а также в обеспечении избирательного возбуждения определенных рабочих органов.
Парабиотическое торможениеразвивается при патологических состояниях, когда лабильность структур центральной нервной системы снижается или происходит очень массивное одновременное возбуждение большого числа афферентных путей, как, например, при травматическом шоке.
Некоторые исследователи выделяют еще один вид торможения - торможение вслед за возбуждением. Оно развивается в нейронах после окончания возбуждения в результате сильной следовой гиперполяризации мембраны (постсинаптической).