

Нейрофизиология / 11 вопрос
.docxПотенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в виде кратковременного изменения мембранного потенциала на небольшом участке возбудимой клетки (нейрона иликардиомиоцита), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, в то время, как в покое она заряжена положительно. Потенциал действия является физиологической основой нервного импульса.
Благодаря работе «калий-натриевого насоса» концентрация ионов натрия в цитоплазме клетки очень мала по сравнению с окружающей средой. При проведении потенциала действия открываются потенциал-зависимые натриевые каналы и положительно заряженные ионы натрия поступают в цитоплазму по градиенту концентрации, пока он не будет уравновешен положительным электрическим зарядом. Вслед за этим потенциал-зависимые каналы инактивируются и отрицательный потенциал покоя восстанавливается за счёт диффузии в клетку отрицательно заряженных ионов хлора, концентрация которых в окружающей среде также значительно выше внутриклеточной.
Фазы потенциала действия
-
Предспайк — процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ).
-
Пиковый потенциал, или спайк, состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны).
-
Отрицательный следовой потенциал — увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация).
-
Положительный следовой потенциал — от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация).
Общие положения
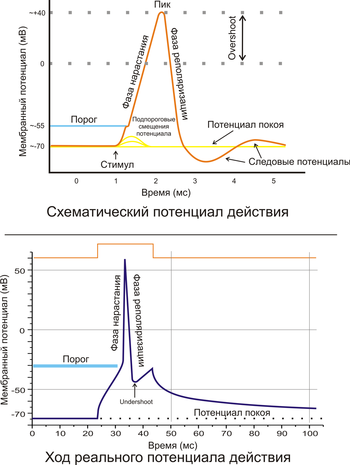
Рис. 2. A. Схематичное изображение идеализированного потенциала действия. B. Реальный потенциал действия пирамидного нейрона гиппокампакрысы. Форма реального потенциала действия обычно отличается от идеализированной.
Поляризация мембраны живой клетки обусловлена отличием ионного состава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемую потенциалом покоя. Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет иметь отрицательное значение (порядка −70 — −90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся и катионы, и анионы. Снаружи — на порядок больше ионов натрия, кальция и хлора, внутри — ионов калия и отрицательно заряженных белковых молекул, аминокислот, органических кислот, фосфатов, сульфатов. Надо понимать, что речь идёт именно о заряде поверхности мембраны — в целом среда и внутри, и снаружи клетки заряжена нейтрально.
Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток, подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий черезсинапс или путём диффузной передачи через межклеточную среду. Смещение мембранного потенциала может происходить в отрицательную (гиперполяризация) или положительную (деполяризация) сторону.
В нервной ткани потенциал действия, как правило, возникает при деполяризации — если деполяризация мембраны нейрона достигает некоторого порогового уровня или превышает его, клетка возбуждается, и от её тела к аксонам и дендритам распространяется волна электрического сигнала. (В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы, которые сильно отличаются от потенциала действия по своей природе — например, они не подчиняются принципу «всё или ничего». Эти потенциалы преобразуются в потенциал действия на особом участке мембраны — аксонном холмике, так что потенциал действия не распространяется на дендриты).
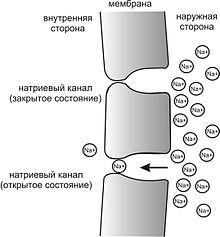
Рис. 3. Простейшая схема, демонстрирующая мембрану с двумя натриевыми каналами в открытом и закрытом состоянии, соответственно
Это обусловлено тем, что на мембране клетки находятся ионные каналы — белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и наоборот. Большинство каналов ионоспецифичны — натриевый канал пропускает практически только ионы натрия и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количество потенциал-зависимых ионных каналов, способных быстро реагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечивается градиентомконцентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны (см. Рис. 2). Поток ионов натрия вызывает ещё бо́льшее и очень быстрое изменение мембранного потенциала, которое и называют потенциалом действия (в специальной литературе обозначается ПД).
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой же амплитуды, как и стимул, превышающий пороговый. Это отнюдь не означает, что амплитуда потенциала действия всегда одинакова — один и тот же участок мембраны, находясь в разных состояниях, может генерировать потенциалы действия разной амплитуды.
После возбуждения нейрон на некоторое время оказывается в состоянии абсолютной рефрактерности, когда никакие сигналы не могут его возбудить снова, затем входит в фазу относительной рефрактерности, когда его могут возбудить исключительно сильные сигналы (при этом амплитуда ПД будет ниже, чем обычно). Рефрактерный период возникает из-за инактивации быстрого натриевого тока, то есть инактивации натриевых каналов (см. ниже).
Распространение потенциала действия
По немиелинизированным волокнам
По немиелинизированному волокну потенциал действия распространяется непрерывно. Проведение нервного импульса начинается с распространениемэлектрического поля. Возникший потенциал действия за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые потенциалы. Сам потенциал действия не перемещается, он исчезает там же, где возник. Главную роль в возникновении нового потенциал действия играет предыдущий.
Если внутриклеточным электродом раздражать аксон посередине, то потенциал действия будет распространяться в обоих направлениях. Обычно же потенциал действия распространяется по аксону в одном направлении (от тела нейрона к нервным окончаниям), хотя деполяризация мембраны происходит по обе стороны от участка, где в данный момент возник потенциал. Одностороннее проведение потенциала действия обеспечивается свойствами натриевых каналов — после открытия они на некоторое время инактивируются и не могут открыться ни при каких значениях мембранного потенциала (свойство рефрактерности). Поэтому на ближнем к телу клетки участке, где до этого уже «прошел» потенциал действия, он не возникает.
При прочих равных условиях распространение потенциала действия по аксону происходит тем быстрее, чем больше диаметр волокна. По гигантским аксонам кальмара потенциал действия может распространяться почти с такой же скоростью, как и по миелинизированным волокнам позвоночных (около 100 м/c).
По миелинизированным волокнам
По миелинизированному волокну потенциал действия распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; здесь их плотность в 100 раз больше, чем в мембранах безмиелиновых волокон. В области миелиновых муфт потенциалзависимых каналов почти нет. Потенциал действия, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до критического уровня, что приводит к возникновению в них новых потенциал действия, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает 2-й, 3-ий, 4-й и даже 5-й, посколькуэлектроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля.
«Скачкообразное распространение» увеличивает скорость распространения потенциала действия по миелинизированным волокнам по сравнению с немиелинизированными. Кроме того, миелинизированные волокна толще, а электрическое сопротивление более толстых волокон меньше, что тоже увеличивает скорость проведения импульса по миелинизированным волокнам. Другим преимуществом сальтаторного проведения является его экономичность в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых меньше 1 % мембраны, и, следовательно, необходимо значительно меньше энергии для восстановления трансмембранных градиентов Na+ и K+, расходующихся в результате возникновения потенциал действия, что может иметь значение при высокой частоте разрядов, идущих по нервному волокну.
Чтобы представить, насколько эффективно может быть увеличена скорость проведения за счёт миелиновой оболочки, достаточно сравнить скорость распространения импульса по немиелинизированным и миелинизированным участкам нервной системы человека. При диаметре волокна около 2 µм и отсутствии миелиновой оболочки скорость проведения будет составлять ~1 м/с, а при наличии даже слабой миелинизации при том же диаметре волокна — 15—20 м/с. В волокнах большего диаметра, обладающих толстой миелинововой оболочкой, скорость проведения может достигать 120 м/с.
Скорость распространения потенциала действия по мембране отдельно взятого нервного волокна не является постоянной величиной — в зависимости от различных условий, эта скорость может очень значительно уменьшаться и, соответственно, увеличиваться, возвращаясь к некоему исходному уровню.
Активные свойства мембраны

Схема строения мембраны клетки.
Активные свойства мембраны, обеспечивающие возникновение потенциала действия, основываются главным образом на поведении потенциалзависимых натриевых (Na+) и калиевых (K+) каналов. Начальная фаза ПД формируется входящим натриевым током, позже открываются калиевые каналы и выходящий K+-ток возвращает потенциал мембраны к исходному уровню. Исходную концентрацию ионов затем восстанавливает натрий-калиевый насос.
По ходу ПД каналы переходят из состояния в состояние: у Na+ каналов основных состояний три — закрытое, открытое и инактивированное (в реальности дело сложнее, но этих трёх достаточно для описания), у K+ каналов два — закрытое и открытое.
Поведение каналов, участвующих в формировании ПД, описывается через проводимость и высчиляется черезкоэффициенты переноса (трансфера).
Коэффициенты переноса были выведены Ходжкином и Хаксли.[1][2]
Потенциал действия (ПД) - это кратковременные высоко амплитуды и изменения МПС, возникающих при возбуждении. Основной причиной ПД является изменение проницаемости мембраны для ионов. Рассмотрим развитие ПД на примере нервного волокна. Регистрировать ПД можно при введении одного из электродов в волокно или располагая оба электрода на его поверхности. Проследим процесс формирования ПД при внутриклеточном методе. 1. В состоянии покоя мембрана поляризована и МПС равен 90 мВ. 2. Как только начинается возбуждение, величина этого потенциала уменьшается (это уменьшение называется деполяризацией). В ряде случаев потенциал сторон мембраны меняется на противоположный (так называемый овершут). Это первая стадия ПД - деполяризация. 3. Стадия реполяризации, при которой величина разности потенциалов падает почти до первоначального уровня. Эти две фазы в пиком ПД. 4. После пика наблюдаются следовые потенциалы - следовая деполяризация и следовая гиперполяризация (гиперполяризация - увеличение разности потенциалов между сторонами мембраны). Например, было 90 мВ, а становится 100 мВ. Развивается ПД очень быстро - за несколько миллисекунд. Параметры ПД: 1) переменный характер, поскольку меняется направление движения тока, 2) величина, которая благодаря овершута может превышать МПС; 3) время, в течение которого развиваются ПД и отдельные его стадии - деполяризация, реполяризация, следовая гиперполяризация. Как формируется ПД. В состоянии покоя «ворота» потенциалзависимых Na +-каналов закрыты. Закрытые также «ворота» потенциалзависимых К +-каналов. 1. Во время фазы деполяризации происходит активизация Na +-Kaналив. При этом изменяется конформационный состояние белков, входящих в состав «ворот». Эти «ворота» открываются, и проницаемость мембраны для Na + увеличивается в несколько тысяч раз. Na + лавообразные входит в волокно нерва. В настоящее время К +-каналы открываются очень медленно. Так, в волокно поступает значительно больше Na +, чем выводится из него К +. 2. Реполяризация характеризуется закрытием Na +-каналов. «Ворота» на внутренней поверхности мембраны закрываются - наблюдается инактивация каналов под воздействием электрических потенциалов. Инактивация происходит медленнее, чем активация. В настоящее время ускоряется активация К +-каналов и растет диффузия К + наружу. Таким образом, деполяризация связана преимущественно со входом Na + в волокно, а реполяризация - с выходом из него К +. Соотношение между входом Na + и выходом К + изменяется в процессе раз «витке ПД: в начале ПД входит Na + в несколько тысяч раз больше, чем получается К +, а затем выходит более К +, чем входит Na +. Причиной следовых потенциалов дальнейшие изменения соотношения между этими двумя процессами. Во время следовой гиперполяризации много К +-каналов еще остаются открытыми и К + продолжает выходить наружу. Восстановление ионных градиентов после ПД. Одиночные ПД изменяют разность концентраций ионов в нервном волокне и за его пределами очень мало. Но в тех случаях, когда проходит значительное количество импульсов, эта разница может быть весьма существенной. Восстановление ионных градиентов происходит тогда за счет усиления работы Na + / K +-HacociB - в большей степени нарушается этот градиент, тем интенсивнее работают насосы. При этом используется энергия АТФ. Часть ее выделяется в виде тепла, поэтому в этих случаях наблюдается кратковременное повышение температуры волокна. Условия необходимые для возникновения ПД. ПД возникает лишь при определенных условиях. Раздражители, действующие на волокну, могут быть разными. Чаще используется постоянный электрический ток. Он легко дозируется, мало травмирует ткань и ближайший тех раздражителей, которые существуют в живых организмах. При каких условиях постоянный ток может зумовитК появление ПД? Ток должен быть достаточно сильным, действовать определенное время, его нарастание должно быть быстрым. Наконец, имеет значение и направление тока (действие анода или катода). В зависимости от силы различают подпороговый (недостаточный для возникновения возбуждения), пороговый (достаточный) и надпороговый (чрезмерный) ток. Несмотря на то что подпороговый ток не вызывает возбуждение, он все же деполяризует мембрану, и эта деполяризация тем больше, чем выше его напряжение. Деполяризация, развивающийся при этом, называется локальным ответом и является видом местного возбуждения. Оно характеризуется тем, что не распространяется, величина его зависит от силы раздражения (закрном силовых отношений: чем больше сила раздражения, тем активнее ответ). При локальной ответа возбудимость ткани повышается. Возбудимость - это способность отвечать на раздражение и переходить в состояние возбуждения. Если сила раздражителя достаточное (пороговая), то деполяризация достигает определенной величины, называется критическим уровнем деполяризации (Ек). Для нервного волокна, покрытого миелином, Ек составляет около 65 мВ. Таким образом, разница между МПС (Е0), равный в данном случае 90 мВ, и Ек составляет 25 мВ. Эта величина (ДЕ = Е0-Ек) очень важна для характеристики возбудимости ткани. Когда Е0 увеличивается при деполяризации, возбудимость выше и, наоборот, снижение Е0 при гиперполяризации приводит к ее уменьшению. ГДЕ может зависеть не только от величины Е0, но и от критического уровня деполяризации (Ек). При пороговой силе раздражителя возникает ПД. Это уже не местное возбуждение, оно способно распространяться на дальние расстояния, подлежит закону «все или ничего» (при увеличении силы раздражителя амплитуда ПД не растет). Возбудимость при развитии ПД отсутствует или значительно снижена. ПД является одним из показателей возбуждения - активного физиологического процесса, которым живые клетки (нервные, мышечные, железистые) отвечают на раздражение. Во время возбуждения меняются метаболизм, температура клеток, нарушается ионная равновесие между цитоплазмой и внешней средой, происходит ряд других процессов. Кроме силы постоянного тока, возникновения ПД зависит также от длительности его действия. Существует обратная пропорциональная зависимость между силой тока и длительностью его действия. Подпороговый ток даже при очень длительном воздействии не приведет возбуждения. Надпороговый ток при слишком короткой действия также не приведет к возбуждению. Для возникновения возбуждения требуется также определенная скорость (крутизна) нарастание силы тока. Если повышать силу тока очень медленно, то при этом будет меняться Ек и Е0 может не достичь его уровня. Имеет значение и направление тока: ПД возникает при замыкании тока только тогда, когда катод помещается на внешней поверхности мембраны, а анод - в клетке или волокне. При прохождении тока изменяется МП. Если на поверхности лежит катод, то развивается деполяризация (возбудимость повышается), а если анод - гиперполяризация (возбудимость снижается). Знание механизмов действия электрического тока на живые объекты крайне необходимо для разработки и применения в клинике методов физиотерапии (диатермия, УВЧ, гипергидроз и др.).. Изменение возбудимости при ПД. При локальной ответа возбудимость повышается (ДЭ уменьшается). Изменения возбудимости во время самого ПД можно заметить, если раздражать повторно в разные стадии развития ПД. Оказывается, что во время пика даже очень сильное повторное раздражение остается без ответа (период абсолютной рефрактерности). Затем возбудимость постепенно нормализуется, но она все же ниже, чем начальная (период относительной рефрактерности). При выраженной следовой деполяризации возбудимость выше, чем первоначальная, а при положительной следового потенциала возбудимость снова снижается. Абсолютная рефрактерность объясняется инактивизациею Na + каналов и повышением проводимости К + - каналов. При относительной рефрактерности вновь активизируются Na + - каналы и снижается правиднисть К + - каналов. Двухфазный характер ПД. Обычно в условиях, когда микроэлектрод содержится внутри клетки или волокна, наблюдается однофазный ПД. Иная картина бывает в тех случаях, когда оба электрода лежат на внешней поверхности мембраны - биполярная регистрация. Возбуждение, которое представляет собой волну электроотрицательности, перемещаясь по мембране, доходит сначала до одного электрода, затем помещается между электродами, наконец достигает второго электрода, а затем распространяется дальше. В этих условиях ПД имеет двухфазный характер. Регистрация ПД широко используется в клинике для диагностики заболеваний сердца, мозга, опорно-двигательного аппарата, желудка и др.
Потенциал действия. Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя, в этом участке возникает возбуждение, одним из наиболее важных проявлений которого служит быстрое колебание МП, называемое потенциалом действия (ПД)
При внутриклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень короткий интервал, измеряемый тысячными долями секунды, становится заряженным электроотрицательно по отношению к соседнему, покоящемуся участку, т.е. при возбуждении происходит т.н. "перезарядка мембраны". Точные измерения показали, что амплитуда ПД на 30-50 мв превышает величину МП. Причина этого состоит в том, что при возбуждении происходит не просто исчезновение ПП, а возникает разность потенциалов обратного знака, в результате чего наружная поверхность мембраны становится заряженной отрицательно по отношению у ее внутренней стороне.
В ПД принято различать его пик (т.н. спайк - spike) и следовые потенциалы. Пик ПД имеет восходящую и нисходящую фазы. Перед восходящей фазой регистрируется более или менее выраженный т.н. местный потенциал , или локальный ответ. Поскольку во время восходящей фазы исчезает исходная поляризация мембраны, ее называют фазой деполяризации; соответственно нисходящую фазу, в течение которой поляризация мембраны возвращается к исходному уровню, называется фазой реполяризации. Продолжительность пика ПД в нервных и скелетных мышечных волокнах варьирует в пределах 0,4-5,0 мсек. При этом фаза реполяризации всегда продолжительнее.
Кроме
пика, в ПД различают два следовых
потенциала - следовую деполяризацию
и следовую гиперполяризацию. Амплитуда
этих потенциалов не превышает нескольких
милливольт, а длительность варьирует
от нескольких десятков до сотен
миллисекунд. Следовые потенциалы связаны
с восстановительными процессами,
развивающимися в мышцах и нерве после
окончания возбуждения.Следовые потенциалы
непостоянны, и в разных тканях могут
проявляться только следовой деполяризацией
или только следовой гиперполяризацией,
последовательность их проявления тоже
может быть различной.
Причиной возникновения ПД является изменение ионной проницаемости мембраны. В состоянии покоя, как уже говорилось, проницаемость мембраны для К+ превышает натриевую проницаемость. Вследствие этого поток положительно заряженных ионов из протоплазмы наружу превышает противоположный поток Na+. Поэтому мембрана в покое снаружи заряжена положительно.
При действии на клетку раздражителя проницаемость мембраны для ионов Na+ резко повышается, и в конечном итоге становится примерно в 20 раз больше проницаемости для К+ Поэтому поток ионов Na+ в клетку начинает значительно превышать направленный наружу поток К+. Ток Na+ достигает величины +150 мв. Одновременно несколько уменьшается выход К+ из клетки. Все это приводит к извращению (реверсии) МП, и наружная поверхность мембраны становится заряженной электро отрицательно по отношению к внутренней поверхности. Указанный сдвиг и регистрируется в виде восходящей ветви пика ПД (фаза деполяризации).
Повышение проницаемости мембраны для ионов Na+ продолжается в нервных клетках очень короткое время. Связано оно с кратковременным открытием т.н. Na+-каналов (точнее, заслонок М в этих каналах), которое затем сменяется срочным закрытием Na+-пор с помощью т.н. Н-ворот. Этот процесс называется натриевой инактивацией. В результате поток Na в клетку прекращается.
В результате Na-инактивации и одновременного увеличения К- проницаемости происходит усиленный выход положительных ионов К+ из протоплазмы во внешний раствор. В итоге этих двух процессов происходит восстановление поляризованного состояния мембраны (реполяризация) , и наружная ее поверхность вновь приобретает положительный заряд. В дальнейшем происходят процессы восстановления нормального ионного состава клетки и необходимого градиента концентрации ионов за счет активизации деятельности Na-К-насоса.
Условия возникновения возбуждения. Для возникновения ПД необходимо, чтобы под влиянием какого-либо раздражителя произошло повышение ионной проницаемости мембраны возбудимой клетки. Однако, возбуждение возможно лишь при условии, если действующий на мембрану агент имеет некоторую минимальную (пороговую) величину, способную изменить мембранный потенциал (МП, или Ео ) до некоторого критического уровня (Ек, критический уровень деполяризации). Стимулы, сила которых ниже пороговой величины, называются подпороговыми, выше - надпороговыми. Показано, что пороговая сила, необходимая для возникновения возбуждения при внутриклеточном микроэлектроде равна 10 -7 - 10-9 А.
Таким образом, главным условием для возникновения ПД является следующее: мембранный потенциал должен стать равным или меньше критического уровня деполяризации ( Ео <= Eк)
Закон "все или ничего". ПД подчиняется закону "все или ничего". При изучении зависимости эффектов раздражения от силы приложенного стимула был установлен т.н. закон "все или ничего". Согласно этому закону, подпороговые раздражения не вызывают возбуждения ("ничего"), при пороговых же стимулах возбуждение сразу приобретает максимальную величину ("все"), и уже не возрастает при дальнейшем усилении раздражителя.
Эта закономерность первоначально была открыта Боудичем при исследовании сердца, а в дальнейшем подтверждена и на других возбудимых тканях. Долгое время закон "все или ничего" неправильно интерпретировался как общий принцип реагирования возбудимых тканей. Предполагали, что "ничего" означает полное отсутствие ответа на под пороговый стимул, а "все" рассматривалось как проявление полного исчерпания возбудимым субстратом его потенциальных возможностей. Дальнейшие исследования, в особенности микроэлектродные, показали, что эта точка зрения не соответствует действительности. Выяснилось, что при под пороговых силах возникает местное не распространяющееся возбуждение (локальный ответ). Вместе с тем оказалось, что "все" также не характеризует того максимума, которого может достигнуть ПД. В живой клетке существуют процессы, активно приостанавливающие деполяризацию мембраны. Если каким-либо воздействием на нервное волокно, например, наркотиками, ядами, ослабить входящий Na-ток, обеспечивающий генерацию ПД, то он перестает подчиняться правилу "все или ничего" - его амплитуда начинает градуально зависеть от силы стимула. Поэтому "все или ничего" рассматривается сейчас не как всеобщий закон реагирования возбудимого субстрата на раздражитель, а лишь как правило, характеризующее особенности возникновения ПД в данных конкретных условиях.