

|
Понятие синапса. Виды синапсов | ||||||||||||||||||||||||||||||
|
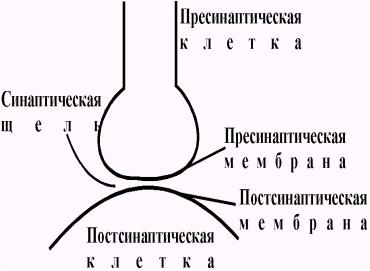
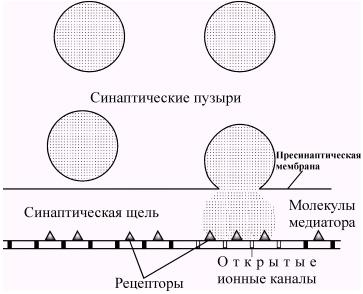
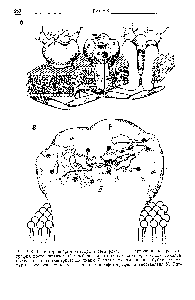
Термин синапс (от греческого sy'napsys - соединение, связь) ввел И. Шеррингтон в 1897 году. В настоящее время синапсами называют специализированные функциональные контакты между возбудимыми клетками (нервными, мышечными, секреторными), служащие для передачи и преобразования нервных импульсов. По характеру контактирующих поверхностей различают: аксо-аксональные, аксо-дендритические, аксо-соматические, нервно-мышечные, нейро-капиллярные синапсы. Электронно-микроскопические исследования выявили, что синапсы имеют три основных элемента: пресинаптическую мембрану, постсинаптическую мембрану и синаптическую щель (рис. 37).
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 37. Основные элементы синапса.
| ||||||||||||||||||||||||||||||
|
Передача информации через синапс может осуществляться химическим или электрическим путем. Смешанные синапсы сочетают химические и электрические механизмы передачи. В литературе на основании способа передачи информации принято выделять три группы синапсов - химические, электрические и смешанные.
| ||||||||||||||||||||||||||||||
|
Строение химических синапсов | ||||||||||||||||||||||||||||||
|
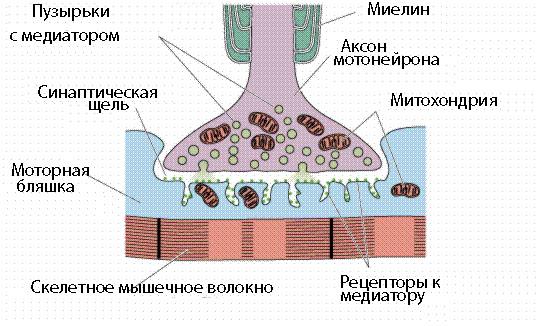
Передача информации в химических синапсах осуществляется через синаптическую щель - область внеклеточного пространства шириной 10-50 нм, разделяющую мембраны пре- и постсинаптических клеток. В пресинаптическом окончании содержатся синаптические везикулы (рис. 38) - мембранные пузырьки диаметром около 50 нм., в каждом из которых заключено 1х104 - 5х104 молекул медиатора. Общее количество таких пузырьков в пресинаптических окончаниях составляет несколько тысяч. Цитоплазма синаптической бляшки содержит митохондрии, гладкий эндоплазматический ретикулум, микрофиламенты (рис. 39).
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 38. Строение химического синапса
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 39. Схема нервно-мышечного синапса
| ||||||||||||||||||||||||||||||
|
Синаптическая щель заполнена мукополисахаридом, "склеивающим" пре- и постсинаптическую мембраны. | ||||||||||||||||||||||||||||||
|
Постсинаптическая мембрана содержит крупные белковые молекулы, выполняющие функции рецепторов, чувствительных к медиатору, а также многочисленные каналы и поры, через которые в постсинаптический нейрон могут поступать ионы.
| ||||||||||||||||||||||||||||||
|
Передача информации в химических синапсах | ||||||||||||||||||||||||||||||
|
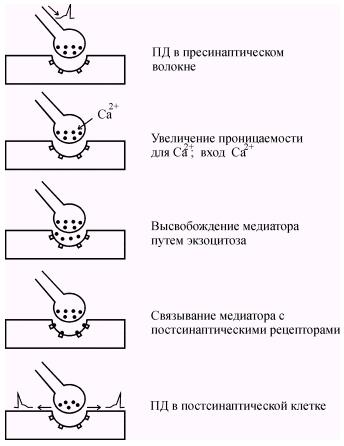
При поступлении потенциала действия к пресинаптическому окончанию происходит деполяризация пресинаптической мембраны и повышается ее проницаемость для ионов Ca2+ (рис. 40). Повышение концентрации ионов Ca2+ в цитоплазме синаптической бляшки инициирует экзоцитоз везикул наполненных медиатором (рис. 41). | ||||||||||||||||||||||||||||||
|
Содержимое везикул высвобождается в синаптическую щель, и часть молекул медиатора диффундирует, связываясь с рецепторными молекулами постсинаптической мембраны. В среднем каждая везикула содержит около 3000 молекул медиатора, а диффузия медиатора до постсинаптической мембраны занимает около 0,5 мс.
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 40. Последовательность событий, происходящих в химическом синапсе от момента возбуждения пресинаптического окончания до возникновения ПД в постсинаптической мембране.
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 41. Экзоцитоз синаптических пузырьков с медиатором. Пузырьки сливаются с плазматической мембраной и выбрасывают свое содержимое в синаптическую щель. Медиатор диффундирует к постсинаптической мембране и связывается с расположенными на ней рецепторами. (Экклз, 1965).
| ||||||||||||||||||||||||||||||
|
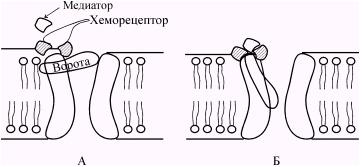
При связывании молекул медиатора с рецептором его конфигурация изменяется, что приводит к открытию ионных каналов (рис. 42) и поступлению через постсинаптическую мембрану в клетку ионов, вызывающих развитие потенциала концевой пластинки (ПКП). ПКП есть результат местного изменения проницаемости постсинаптической мембраны для ионов Na+ и К+. Но ПКП не активирует другие хемовозбудимые каналы постсинаптической мембраны и его величина зависит от концентрации медиатора, действующего на мембрану: чем больше концентрация медиатора, тем выше (до определенного предела) ПКП. Таким образом, ПКП в отличие от потенциала действия градуален. В этом отношении он схож с локальным ответом, хотя механизм его возникновения иной. При достижении ПКП некоторой пороговой величины возникают местные токи между участком деполяризованной постсинаптической мембраны и соседними с ней участками электровозбудимой мембраны, что вызывает генерацию потенциала действия.
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 42. Строение и работа хемовозбудимого ионного канала. Канал образован макромолекулой белка, погруженной в липидный би слой мембраны. До взаимодействия молекулы медиатора с рецептором ворота закрыты (А). Они открываются при связывании медиатора с рецептором (Б). ( По Ходорову Б.И.).
| ||||||||||||||||||||||||||||||
|
Таким образом, процесс передачи возбуждения через химический синапс может быть схематически представлен в виде следующей цепи явлений: потенциал действия на пресинаптической мембране поступление ионов Ca2+ внутрь нервного окончания освобождение медиатора диффузия медиатора через синаптическую щель к постсинаптической мембране взаимодействие медиатора с рецептором активация хемовозбудимых каналов постсинаптической мембраны возникновение потенциала концевой пластинки критическая деполяризация постсинаптической электровозбудимой мембраны генерация потенциала действия. | ||||||||||||||||||||||||||||||
|
Химические синапсы имеют два общих свойства: | ||||||||||||||||||||||||||||||
|
1. Возбуждение через химический синапс передается только в одном направлении - от пресинаптической мембраны к постсинаптической мембране (одностороннее проведение). | ||||||||||||||||||||||||||||||
|
2. Возбуждение проводится через синапс значительно медленнее, чем по нервному волокну синаптическая задержка. | ||||||||||||||||||||||||||||||
|
Односторонность проведения обусловлена высвобождением медиатора из пресинаптической мембраны и локализацией рецепторов на постсинаптической мембране. Замедление проведения через синапс (синаптическая задержка) возникает вследствие того, что проведение является многоэтапным процессом (секреция медиатора, диффузия медиатора к постсинаптической мембране, активация хеморецепторов, рост ПКП до пороговой величины) и для протекания каждого из перечисленных этапов требуется время. Кроме этого, наличие относительно широкой синаптической щели препятствует проведению импульса с помощью локальных токов.
| ||||||||||||||||||||||||||||||
|
Химические медиаторы | ||||||||||||||||||||||||||||||
|
Медиаторы (от латинского - mediator - проводник) - биологически активные вещества, посредством которых осуществляются межклеточные взаимодействия в синапсах. | ||||||||||||||||||||||||||||||
|
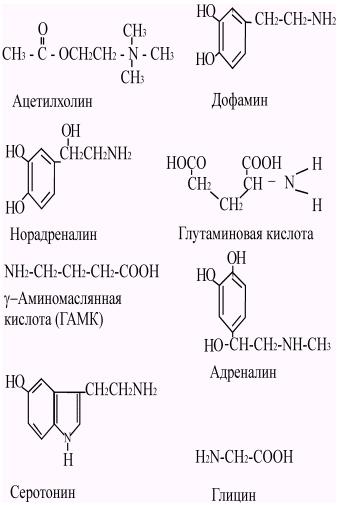
В основном химическими медиаторами являются низкомолекулярные вещества. Однако некоторые высокомолекулярные соединения, такие как полипептиды, могут также выполнить роль химических посредников. В настоящее время известен ряд веществ, играющих роль медиаторов в ЦНС млекопитающих. К ним относятся ацетилхолин, биогенные амины: адреналин, норадреналин, дофамин, серотонин, кислые аминокислоты: глицины, гамма-аминомаслянная кислота (ГАМК), полипептиды: вещество Р, энкефалин, соматостатин и др. (Рис. 43).
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 43. Структурные формулы некоторых медиаторов.
| ||||||||||||||||||||||||||||||
|
Функцию медиаторов могут выполнять и такие соединения как АТФ, гистамин, простагландины. В 1935 году Г.Дейлом было сформулированно правило (принцип Дейла), согласно которому каждая нервная клетка выделяет только один определенный медиатор. Поэтому принято обозначать нейроны по типу медиатора, который выделяется в их окончаниях. Так, нейроны, освобождающие ацетилхолин, называется холинергическими, норадреналин - адренергическими, серотонин - серотонинергическими, амины - аминергическими и т.д.
| ||||||||||||||||||||||||||||||
|
Квантовое выделение медиаторов | ||||||||||||||||||||||||||||||
|
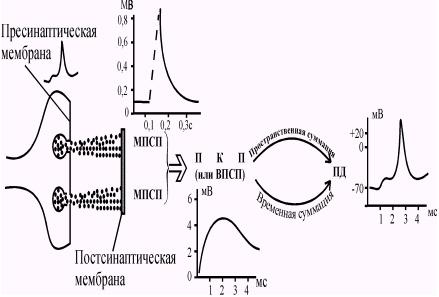
Изучая механизмы нервно-мышечной передачи, Пол Фетт и Бернард Катц в 1952 году зарегистрировали миниатюрные постсинаптические потенциалы (МПСП). МПСП можно зарегистрировать в области постсинаптической мембраны. По мере удаления внутриклеточного регистрирующего электрода от постсинаптической мембраны МПСП постепенно уменьшается. Амплитуда МПСП составляет менее 1 мв. (Рис. 44).
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 44. Миниатюрные постсинаптические потенциалы, записанные в области концевой пластинки волокна скелетной мышцы. Видно, что амплитуда МПСП невелика и постоянна. (По Р.Эккерт ).
| ||||||||||||||||||||||||||||||
|
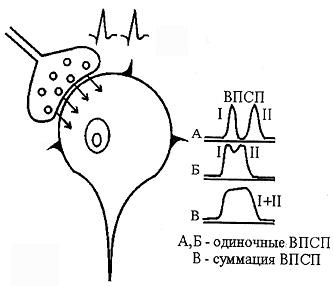
Катц и его сотрудники исследовали, как связаны МПСП с обычными ПКП, возникающими при возбуждении двигательных нервов. Было высказано предложение, что МПСП есть результат выделения "кванта" медиатора, а ПКП складывается в результате суммации многих МПСП. В настоящее время известно, что "квант" медиатора представляет собой "пакет" молекул медиатора в синаптическом пузырьке пресинаптической мембраны. По расчетам, каждый МПСП соответствует выбросу кванта медиатора, состоящего из 10000 - 40000 молекул медиатора, что приводит к активации около 2000 постсинаптическтих ионных каналов. Для возникновения потенциала концевой пластинки (ПКП) или возбуждающего постсинаптического потенциала (ВПСП) необходимо выделение 200-300 квантов медиатора.
| ||||||||||||||||||||||||||||||
|
Генерация потенциала действия | ||||||||||||||||||||||||||||||
|
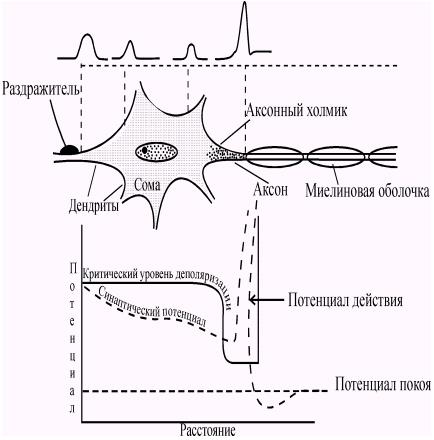
Миниатюрный постсинаптический потенциал, потенциал концевой пластинки и возбуждающий постсинаптический потенциал являются локальными процессами. Они не могут распространяться и, следовательно, не могут обеспечивать передачу информации между клетками. Участком генерации потенциалов действия в мотонейроне служит начальный сегмент аксона, расположенный непосредственно за аксонным холмиком (рис. 45). Этот участок наиболее чувствителен к деполяризации и обладает более низким критическим уровнем деполяризации, чем тело и дендриты нейрона. Поэтому именно в области аксонного холмика возникают потенциалы действия. Для того, чтобы вызвать возбуждение, ПКП (или ВПСП) должны достигнуть в области аксонного холмика некоторого порогового уровня (рис. 46).
Рис. 46. Пространственное затухание ВПСП и генерация потенциала действия. Возбуждающие синаптические потенциалы, возникающие в дендрите, по мере распространения по нейрону затухают. Порог генерации ПД (критический уровень деполяризации) зависит от плотности натриевых каналов (черные точки). Хотя синаптический потенциал (он изображен в верхней части рисунка) и затухает по мере распространения от дендрита к аксону, ПД все же возникает в области аксонного холмика. Именно здесь плотность натриевых каналов наиболее высока, а пороговый уровень деполяризации наиболее низок. (Р. Эккерт).
| ||||||||||||||||||||||||||||||
|
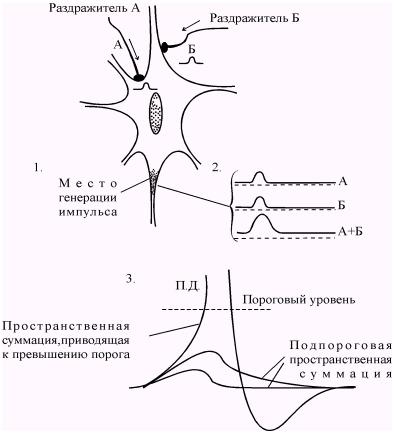
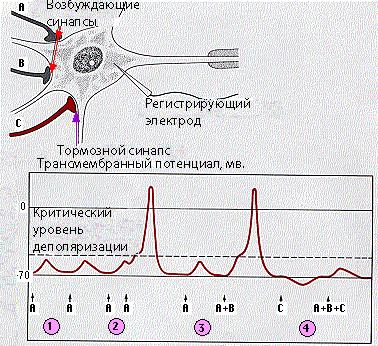
Важное значение для возникновения потенциала действия в нервной клетке имеет суммация возбуждающих синаптических влияний, так как деполяризации, создаваемой одним синапсом, часто бывает недостаточно для достижения порогового уровня и генерации потенциала действия. Так, если происходит увеличение ВПСП за счет сложения потенциалов, возникающих благодаря работе разных синапсов, то имеет место пространственная суммация (рис. 48). Критический уровень деполяризации может быть достигнут и благодаря временной суммации (рис. 47).
Рис. 47. Схема сомото-дентритных синапсов, обеспечивающих суммацию возбуждения. Так, если после одного постсинаптического потенциала возникает другой, то второй потенциал "накладывается" на первый, в результате чего образуется суммарный потенциал с большей амплитудой (рис. 49.). | ||||||||||||||||||||||||||||||
|
Чем короче при этом будет интервал между двумя последовательными синаптическими потенциалами, тем выше будет амплитуда суммарного потенциала. В естественных условиях обычно одновременно происходит как пространственная, так и временная суммации. Таким образом, за период между выделением медиатора в синаптическую щель и возникновением потенциала действия на постсинаптической структуре (нейроне, мышце, железе) осуществляется ряд биоэлектрических явлений, последовательность и специфические особенности которых представлены в (табл. 1) и (рис. 51.).
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 48. Пространственная суммация в мотонейроне
Рис 49. Временная суммация. При высокой частоте следования раздражителей возможно «наложение» одного постсинаптического потенциала на другой, в результате чего образуется суммарный потенциал с большей амплитудой.
| ||||||||||||||||||||||||||||||
|
1. Возбуждающие постсинаптические потенциалы, возникающие в двух разных синапсах (А и Б ). | ||||||||||||||||||||||||||||||
|
2. Потенциалы, возникающие на мембране в зоне генерации импульса при раздражении волокна А или Б либо обоих этих волокон одновременно (А+Б). | ||||||||||||||||||||||||||||||
|
3. Для того чтобы потенциал в области аксонного холмика превышал пороговый уровень, необходима пространственная суммация ВСПС, возникающих в нескольких синапсах. ( Р. Эккерт).
| ||||||||||||||||||||||||||||||
|
Кроме возбуждающих синапсов, через которые передается возбуждение, имеются тормозные синапсы, в которых медиаторы (в частности, ГАМК) вызывают торможение на постсинаптической мембране (рис. 50). В таких синапсах возбуждение пресинаптической мембраны приводит к выделению тормозного медиатора, который действуя на постсинаптическую мембрану, обусловливает развитие ТПСП (тормозного постсинаптического потенциала). Механизм его возникновения связан с увеличением проницаемости постсинаптической мембраны для К+ и Cl-, в результате чего происходит ее гиперполяризация. Подробнее механизм торможения будет изложен в следующей лекции.
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 50. Схема пространственной суммации при наличии возбуждающих и тормозящих синапсов.
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
ТАБЛИЦА № 1.
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 51. Последовательность биоэлектрических явлений в химическом синапсе происходящих за время между выделением медиатора и возникновением ПД на постсинаптической структуре.
| ||||||||||||||||||||||||||||||
|
Метаболизм медиаторов | ||||||||||||||||||||||||||||||
|
Ацетилхолин, выделяемый окончаниями холинергических нейронов, гидролизуется до холина и ацетата ферментом ацетилхолинэстеразой. Продукты гидролиза на постсинаптическую мембрану не действуют. Образующийся холин активно поглощается пресинаптической мембраной и взаимодействуя с ацетилкоферментом А, образует новую молекулу ацетилхолина. (Рис. 52.).
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 52. Метаболизм ацетилхолина (АцХ ) в холинэнергическом синапсе. АцХ, поступающий из пресинаптического окончания, гидролизуется в синаптической щели ферментом ацетилхолинестеразой (АцХЭ). Холин поступает в пресинаптическое волокно и используется для синтеза молекул ацетилхолина (Mountcastle, Baldessarini, 1968)
| ||||||||||||||||||||||||||||||
|
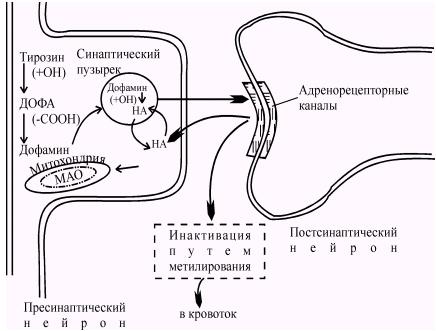
Аналогичный процесс происходит и с другими медиаторами. Другой хорошо изученный медиатор - норадреналин выделяется постганглионарными синаптическими клетками и хромаффинными клетками мозгового слоя надпочечников. Биохимические превращения, которые претерпевает норадреналин в адренергических синапсах, схематично представлен на рисунке 53.
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 53. Биохимические превращения медиатора в адренергическом синапсе. Норадреналин (НА) синтезируется из аминокислоты фенилаланина с образованием промежуточного продукта - тирозина. Образующийся НА запасается в синаптических пузырьках. После высвобождения из синапса часть НА обратно захватывается пресинаптическим волокном, а другая часть инактивируется путем метилирования и удаляется с кровотоком. НА, попавший в цитоплазму пресинаптического окончания, либо захватывается в синаптические пузырьки, либо разрушается моноаминоксидазой (МАО). (Mountcastle, Baldessarini, 1968).
| ||||||||||||||||||||||||||||||
|
Синаптическая модуляция | ||||||||||||||||||||||||||||||
|
Биохимические процессы, осуществляющиеся в синапсе, в значительной степени подвержены влиянию различных факторов - в первую очередь, химических. Так, ацетилхолинэстераза может быть инактивирована некоторыми нервно-паралитическими веществами и инсектицидами. В этом случае ацетилхолин накапливается в синапсах. Это приводит к нарушению реполяризации постсинаптической мембраны и инактивированию холинорецепторов (рис. 54.). В результате нарушается деятельность межнейронных и нервно-мышечных синапсов и быстро наступает гибель организма. Однако в нервной системе образуется большое количество веществ, играющих роль синаптических модуляторов - веществ, влияющих на синаптическое проведение.
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 54. Влияние ингибитора холинэстеразы (неостигмина) на длительность постсинаптического потенциала одиночного мышечного волокна.а - до применения неостигмина; б - после применения неостигмина.(По Б.И. Ходорову).
| ||||||||||||||||||||||||||||||
|
По химической природе эти вещества - пептиды, однако часто их называют нейропептидами, хотя далеко не все из них образуются в нервной системе. Так, ряд веществ синтезируется в эндокринных клетках кишечника, а некоторые нейропептиды первоначально были обнаружены во внутренних органах. Наиболее известными веществами такого рода являются гормоны желудочно-кишечного тракта - глюкагон, гастрин, холецистокинин, вещество Р, желудочный ингибирующий пептид (ЖИП). | ||||||||||||||||||||||||||||||
|
Значительный интерес исследователей вызывают две группы нейропептидов - эндорфины и энкефалины. Эти вещества обладают анальгетическими (уменьшающими боль), галюциногенными, а также некоторыми другими свойствами (вызывают ощущение удовлетворения и эйфории, их активация учащает пульс и повышает температуру тела). Обезболивающий эффект этих соединений может быть связан с тем, что эти нейропептиды препятствуют выделению медиаторов из некоторых нервных окончаний. С такой точкой зрения хорошо согласуется тот факт, что энкефалины и эндорфины присутствуют в задних рогах спинного мозга, т.е. в той области, где в спинной мозг входят чувствительные пути. Болевые ощущения могут уменьшаться в результате выделения нейропептидов, нарушающих синаптическое проведение в эфферентных путях, передающи болевые сигналы. Содержание эндорфинов и энкефалинов непостоянно: например, во время приема пищи, болевых воздействиях, прослушивания приятной музыки их выделение увеличивается. Тем самым организм оберегает себя от чрезмерных болевых ощущений и одаривает за биологически полезные действия.Благодаря таким свойствам, а также тому, что эти нейропептиды связываются в нервной системе с теми же рецепторами, что и опиаты (опий и его производные), они называются эндогенными опиоидами. В настоящее время известно, что на поверхности мембраны некоторых нейронов имеются опиоидные рецепторы с которыми в естественных условиях и связываются вырабатываемые нервной системой, энкефалины и эндорфины. Но при употреблении наркотических опиатов - алкалоидных веществ, выделяемых из растений, опиаты связываются с опиоидными рецепторами, вызывая их неестественно мощную стимуляцию. Это вызывает чрезвычайно приятные субъективные ощущения. При неоднократном применении опиоидов возникают компенсаторные изменения метаболизма нервных клеток, и тогда после их отмены состояние нервной системы становится таким, что больной без введения очередной дозы наркотика испытывает чрезвычайный дискомфорт (абстинентный синдром). Подобная метаболическая зависимость называется пристрастием. | ||||||||||||||||||||||||||||||
|
При изучении опиоидных рецепторов весьма полезным оказалось вещество налоксон - конкурентный блокатор этих рецепторов. Поскольку налоксон препятствует связыванию опиатов с клетками-мишенями, с его помощью можно определить, вызвана ли та или иная реакция возбуждением таких рецепторов. Было обнаружено, например, что налоксон в значительной степени снимает анальгетический эффект плацебо (нейтрального вещества, которое дают больным, уверяя их, что оно снимет у них боль). Вероятно, вера в лекарство (или другое средство лечения), которое должно снять боль, приводит к выбросу опиоидных пептидов; возможно, в этом и состоит фармакологический механизм действия плацебо. Налоксон снимает также обезболивающий эффект иглоукалывания. Отсюда был сделан вывод, что при иглоукалывании из ЦНС выбрасываются естественные опиоидные пептиды. | ||||||||||||||||||||||||||||||
|
Таким образом, эффективность синаптической передачи может быть существенно изменена под влиянием веществ (модуляторов), не участвующих непосредственно в передаче информации.
| ||||||||||||||||||||||||||||||
|
Особенности строения и функционирования электрических синапсов | ||||||||||||||||||||||||||||||
|
Электрические синапсы широко распространены в нервной системе беспозвоночных, а у млекопитающих встречаются крайне редко. Вместе с тем электрические синапсы у высших животных широко распространены в сердечной мышце, гладкой мускулатуре внутренних органов печени, эпителиальной и железистых тканях. | ||||||||||||||||||||||||||||||
|
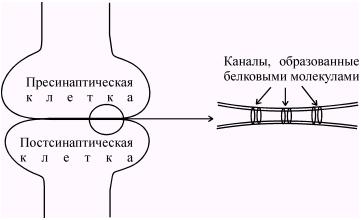
Ширина синаптической щели в электрических синапсах составляет всего 2-4 нм, что значительно меньше чем в химических синапсах. Важной особенностью электрических синапсов является наличие между пресинаптической и постсинаптической мембранах своеобразных мостиков, образованных белковыми молекулами. Они представляют собой каналы шириной 1-2 нм (рис. 55.).
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 55. Структура электрического синапса. Характерные особенности: узкая (2-4 нм) синаптическая щель и наличие каналов, образованных белковыми молекулами.
| ||||||||||||||||||||||||||||||
|
Благодаря наличию каналов, размеры которых позволяют переходить из клетки в клетку неорганическим ионам и даже небольшим молекулам, электрическое сопротивление такого синапса, получившего название щелевого или высокопроницаемого контакта оказывается очень низким. Такие условия позволяют пресинаптическому току распространятся на постсинаптическую клетку практически без угасания. Электрический ток течет от возбужденной области к невозбужденной и там вытекает наружу, вызывая ее деполяризацию (рис. 56.).
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 56. Схема передачи возбуждения в химическом (А) и электрическом синапсе (Б). Стрелками показано распространение электрического тока через мембрану пресинаптического окончания и постсинаптическую мембрану на нейрон. (По Б.И. Ходорову).
| ||||||||||||||||||||||||||||||
|
Электрические синапсы обладают рядом специфических функциональных свойств:
| ||||||||||||||||||||||||||||||
|
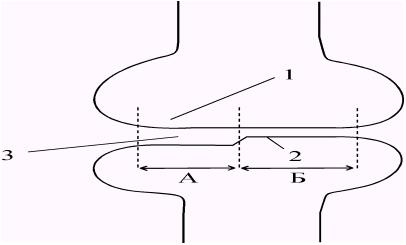
Наряду с химическими и электрическими синапсами между некоторыми нейронами имеются так называемые смешанные синапсы. Их главная особенность заключается в том, что электрическая и химическая передача осуществляется параллельно, поскольку щель между пре- и постсинаптической мембранами имеет участки со структурой химического и электрического синапсов (рис. 57.).
| ||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||
|
Рис. 57. Структура смешанного синапса. А - участок химической передачи. Б - участок электрической передачи. 1. Пресинаптическая мембрана. 2. Постсинаптическая мембрана. 3. Синаптическая щель. |
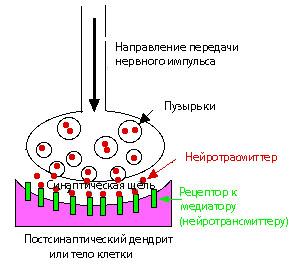





Основные функции синапсов
Значение механизмов функционирования клеток становится понятным при выяснении процессов их взаимодействия, необходимых для обмена информацией. Обмен информацией происходит с помощью нервной системы и в ней самой. Места контактов между нервными клетками (синапсы) играют большую роль при переносе информации. Информация в виде серии потенциалов действия поступает от первого (пресинаптического ) нейрона на второй ( постсинаптический ). Это возможно непосредственно путем формирования локального тока между соседними клетками либо, что гораздо чаще, опосредованно путем химических веществ-переносчиков.
Не вызывает сомнения важность функций клеток для успешной работы всего организма. Однако чтобы организм мог функционировать как единое целое, между его клетками должна осуществляться взаимосвязь - перенос разнообразных химических веществ и информации. В передаче информации участвуют, например, гормоны , доставляемые к клеткам кровью. Но, прежде всего, передача информации осуществляется в нервной системе в виде нервных импульсов. Так, органы чувств получают информацию из окружающего мира, например, в форме звука, света, запаха, и передают ее дальше по соответствующим нервам в мозг. Центральная нервная система , со своей стороны, должна переработать эту информацию и в качестве результата вновь выдать некую информацию на периферию, что образно можно представить в виде определенных приказов на периферические эффекторные органы, такие, например, как мышцы, железы, органы чувств. Это и будет ответом на внешние раздражения.
Проведение информации, например, от рецепторов органа слуха к мозгу включает и ее переработку в ЦНС. Для этого миллионы нервных клеток должны взаимодействовать между собой. Только на основе этой переработки получаемой информации возможно формирование конечного ответа, например, направленные действия или прекращения этих действий, бегство или наступление. Эти два примера свидетельствуют о том, что переработка информации в ЦНС может привести к реакциям, включающим или процессы возбуждения, или процессы торможения. В передаче информации и формировании ответной реакции ЦНС принимают участие и контактные зоны между нервными клетками - синапсы. Помимо синаптических контактов между интернейронами в ЦНС эти процессы осуществляются синаптическими контактами, лежащими на пути передачи эфферентной информации, синапсами междуаксоном и исполнительным нейроном и вне пределов ЦНС (на периферии) между исполнительным нейроном и эффекторным органом. Понятие "синапс" ввел в 1897 г. английский физиолог Ф.Шеррингтон (F.Cherrington). Синапс между аксоном мотонейрона и волокном скелетной мышцы называетсямионевральным синапсом .
Было показано, что при возбуждении нейрон генерирует потенциал действия. Серии потенциалов действия - это носители информации. Задачей синапса является передача этих сигналов от одного нейрона на другой или на эффекторные клетки. Как правило, результатом перекодировки является возникновение потенциалов действия, которые при этом могут подавляться под влиянием других синаптических контактов. В конечном итоге синаптическое проведение вновь приводит к электрическим явлениям. Здесь есть две возможности. Быстрая передача сигналов осуществляется электрическими синапсами , более медленная - химическими , в которых химическое вещество-переносчик берет на себя роль передачи сигнала. Однако и в этом случае имеются две принципиальные возможности. В одном случае химическое вещество-переносчик может вызвать непосредственно электрические явления на мембране соседней клетки, при этом эффект оказывается относительно быстрым. В других случаях это вещество вызывает только цепь дальнейших химических процессов, которые, со своей стороны, ведут к электрическим явлениям на мембране последующего нейрона, что связано с большими временными затратами.
Обычно принята следующая терминология. Если клетка, от которой осуществляется направленное проведение информации, располагается перед синапсом, то она пресинаптическая . Клетка, лежащая после синапса, называется постсинаптической .
Резюме:
- Синапс представляет собой место контакта двух клеток. Информация в виде потенциалов действия поступает от первой клетки, называемой пресинаптической, ко второй, называемой постсинаптической.
- Сигнал через синапс передается электрическим путем посредством возникновения локальных токов между двумя клетками (электрические синапсы), химическим путем, при котором электрический сигнал передается опосредованно при помощи трансмиттера (химические синапсы), и при помощи этих обоих механизмов одновременно (смешанные синапсы).
Синапс электрический
|
Рис. 8.2. Схема никотинового холинэргического синапса. Пресинаптическое нервное окончаниесодержит компоненты для синтеза нейромедиатора (здесь ацетилхолина). После синтеза (I) нейромедиатор упаковывается в пузырьки (везикулы) (II). Эти синаптические везикулы сливаются (возможно, вре.мен-но) с пресинаптической мембраной (1П), и нейромедиатор высвобождается таким путем в синаптическую щель. Он диффундирует к постсинаптической мембране и связывается там со специфическим рецептором(IV). В результате образования нейромедиатор-рецепторного комплекса постсинаптическая мембрана становится проницаемой для катионов (V), т. е. деполяризуется. (Если деполяризация достаточно высока, то появляется потенциал действия, т. е.химический сигнал снова превращается в электрический нервный импульс.) Наконец, медиатор инактивируется , т. е. либо расщепляется ферментом (VI), либо удаляется из синаптической щели посредством особого механизма поглощения. В приведенной схеме только один продукт расщепления медиатора— холин — поглощаетсянервным окончанием (VII) и используется вновь.Базальная мембрана — диффузная структура, идентифицируемая методом электронной микроскопии в синаптической щели (рис. 8.3,а), здесь не показана. |
|
 <=""
img="" style="border: none; display: block;
margin-left: auto; margin-right: auto;">
<=""
img="" style="border: none; display: block;
margin-left: auto; margin-right: auto;">Электрические и химические синапсы [c.188] Свойства электрического синапса [c.189]
|
Рис. 9,8. Рецептор нейромедиатора в мегибране. а — электронная микрофотографияпостсинаптической мембраны никотинового холинэргического синапса, выделенного изэлектрической ткани Torpedo. Напоминающиебублик структуры— это рецепторы (электронная микрофотография предоставлена М, Гир- |
|
 <=""
img="" style="border: none; display: block;
margin-left: auto; margin-right: auto;">
<=""
img="" style="border: none; display: block;
margin-left: auto; margin-right: auto;">Передача сигналов от клетки к клетке. может осуществляться либо путем прямого прохождения потенциалов действия (электрические синапсы), либо с помощью специальных молекул — нейромедиаторов (химические синапсы). В зависимости от своих специфических функций синапсы имеют очень разные структуры. Вхимических синапсах расстояние между клетками составляет - 20—40 нмсинаптическая щель между клетками— это часть межклеточного пространстваона содержит жидкость с низким электрическим сопротивлением, так чтоэлектрический сигнал рассеивается прежде, чем он достигнет следующей клетки.Электрическая передача, напротив, осуществляется только в специализированных структурах — щелевых контактах, где клетки находятся на расстоянии 2 нм и соединяются проводящими канала.ми. В действительности здесь имеется нечто сходное с постулированным ранее синцитием, или многоклеточным цитоплазматическим континуумом. По иронии истории нау- [c.188] Системы пассивного транспорта, называемые далее каналами, не являются единой группой функциональных элементов в мембране. В состоянии покоя каналы закрыты и переходят в проводящее состояние только после их открытия. Открывание, или воротный механизм, запускается электрическим путем, т. е. при изменении мембранного потенциала, или химическим путем — при взаимодействии со специфической молекулой. Химическая природа воротного механизма в тесной связи с биохимией синапса рассмотрена в гл. 8 и 9. Хотелось бы лишь отметить, что воротный механизм также отличается от других транспортныхсистем по своей фармакологии, ионной селективности и кинетике. [c.132] Среди многочисленных примеров, указывающих на значение коммуникативных связей, можно привести явление электрического сопряжения клеток. Обычно мембраны клеток обладают очень высоким электрическим сопротивлением, однако в мембранах соприкасающихся клеток имеются участки с низким сопротивлением — по-видимому, области щелевых контактов [49]. Одна из наиболее совершенных форм коммуникативной связи—это синапс, специализированный контакт между нейронами. Нервный импульс, проходящий по мембране одного нейрона, стимулирует выделение кванта химического вещества (медиатора), который проходит через щель синапса и инициируетвозникновение нервного импульса во втором нейроне. [c.60] Нервное волокно представляет собой сильно вытянутую трубку из студневидного вещества, заполненную солевым раствором одного состава и омываемую солевым раствором другого состава. Эти растворы содержатэлектрически заряженные ионы, по отношению к которым напоминающаямембрану оболочка нерва обладает избирательной проницаемостью. Из-за различия в скоростях диффузии отрицательно и положительно заряженных ионов между внутренней и наружной поверхностью нервного волокнаимеется некоторая разность потенциалов. Если ее мгновенно снизить, то есть вызвать местную деполяризацию, эта деполяризация распространится на соседние участки мембраны, в результате чего по волокну побежит ее волна. Это и есть так называемый спайк-потенциал, или нервный импульс. Мембрана не может быть разряжена частично она деполяризуется полностью на всем пути или не деполяризуется совсем. Кроме того, после прохождения импульса требуется некоторое время для восстановления первоначального потенциала мембраны, причем, до тех пор пока потенциал мембраны не восстановится, нервное волокно не сможет пропустить следующего импульса. Природу возникновения нервного импульса (по закону все или ничего ) и следующего за прохождением импульса рефрактерного периода(или периода возвращения волокна в первоначальное состояние) мы рассмотрим подробнее в последней главе книги. Если возбуждение получено где-то посредине волокна, импульс должен был бы распространяться в обе стороны. Но этого обычно не происходит, так как нервная ткань сконструирована таким образом, чтобы сигнал в любой данный момент шел в каком-то определенном направлении. Для этогонервные волокна соединены между собой в нерве специальными образованиями, синапсами, пропускающими сигналы только в одном направлении. [c.117] Каналы пассивного транспорта ионов, проходящих через возбудимые мембраны, содержат два функциональных компонента воротный механизм иселективный фильтр. Воротный механизм, способный открывать или закрывать канал, может быть активирован электрически путем изменения мембранного потенциала или химически, например в синапсе, связыванием с молекулой нейромедиатора. Селективный фильтр имеет такие размеры и такое строение, которые позволяют пропускать ли- [c.162] Синапсы —это места коммуникации нервных клеток. Химические и электрические синапсы различаются по механизму передачи информации. [c.237] В гл. 1 уже говорилось о то.м, что практически все функции нейронов в большей или меньшей степени обусловлены свойствами мембран. В частности, мембранную природу имеют такие явления как распространение нервных импульсов, их электрическая или химическая передача от клетки к клетке,активный транспорт ионов, клеточное узнавание и развитие синапса, взаимодействие с нейромодуляторами, нейрофармакологическими веществами и нейротоксинами. Такой, несколько односторонний взгляд уточняется в настоящей главе рассмотрением цитоплазмы нейронов. Хотя в основном она сходна с цитоплазмой других клеток — в ней обнаружены те же органеллы (а также синаптические везикулы) и ферменты (и, кроме того, участвующие в метаболизме медиаторы), однако нейрональная цитоплазма адаптирована специфическим образом именно к функциям нейронов. [c.303] ОТ образования микротрубочек или от присутствия медиатора нли Са2+синаптический контакт не обусловлен наличием медиатора, электрической активностью или образованием функциональных рецепторов. Ни одно из исследований, сделанных до сих пор, полностью не отвечает на вопрос о механизме образования, специфичности и стабилизации синапсов и не решает проблемыэтапного образования нейронной сети, ответственной за высшие функции нервной системы. В начале этой главы мы осветили этот вопрос как один изнаиболее важных в нейробиологии, однако подробнее рассмотрим его немного позже. [c.330] Физостигмин сыграл важную роль в истории науки. Он ингибирует фермент холинэстеразу, расщепляющую ацетилхолин (см. разд, 6,2). Благодаря этому последний, как нейромедиатор, долго сохраняется в нервных окончаниях. Это позволило выделить его из них, определить его функцию и вообще развить теорию химической передачи электрического импульса через синапсы нервнойсистемы. [c.531] Основу нервной системы образуют нервные клетки - нейроны, которые связаны между собой синапсами. Благодаря такому строению нервная системаспособна передавать нервные импульсы. Нервный импульс - это электрический сигнал, который двигается по клетке пока не достигнет нервного окончания, где под действием электрического сигнала высвобождаются молекулы, называемые нейромедиаторами. Они и переносят сигнал (информацию) через синапс, достигая другой нервной клетки. [c.54] Биохимических исследований структуры и механизма действияэлектрических синапсов до сих пор не проводилось. Однако щелевыми контактамисвязаны не только нервные клетки, но также и клетки печени, эпителия, мышц имногих других тканей. Из них удалось выделить и охарактеризоватьбиохимическими методами и электронной микроскопией мембранные фрагменты, которые определенно сохраняли зоны межклеточных контактов.Электронные микрофотографии показывают упорядоченные структуры частиц, которые Гудинаф назвал коннексонами [1] и которые образуют каналы между клетками, отстоящими друг от друга на 2 нм. Из этих мембран были выделены два полипептида с М 25 000 и 35 000, названные коннексинами. Возможно, что два коннексона соседних клеток посредством дпме-ризации могут образовать канал(рис. 8.1). Показано, что этот канал пропускает не только ионы щелочных металлов, но п молекулы с М 1000—2000. Таким образом, коннексоны, кромеэлектрического сопряжения, обеспечивают для клеток возможность обмена метаболитами. Проницаемость таких каналов могут регулировать ионыкальция. [c.189] Нейроны представляют собой клетки с длинными отростками, способные про-водить электрические сигналы. Обычно сигналы воспринимаются дендритами ителом клетки, а затем передаются по аксону в виде потенциалов действия. Связь с другими нейронами осуществляется в синапсах, где сигналы передаются с помощью химического вещества-нейромедиатора. Помимо нейронов нервная ткань всегда содержит различные глиальные клетки, которые выполняют поддерживающую функцию. [c.76] Рпс. 19-4. Схема типичного синапса. Электрический сигнал, приходящий в окопчапие аксона клетки , приводит к высвобождению в синаптическую щельхимического посредпика (нейромедиатора), который вызывает электрическое изменение в мембране дендрита клетки В [c.290] В нейрохимическом плане лучше других синапсов изучен электромоторный синапс электрического органа рыб, где нейромедиатором служит АХ. В начале 70-х годов в лаборатории В.Уит-такера в ФРГ впервые удалось выделить изолированную фракцию синаптических пузырьков из электрического органа ската Torpedo marmorata. Именно на этом объекте с помощью биохимических, иммуноцитохимических методов и ядерного магнит- [c.212] Нейроны характеризуются необыкновенно высоким уровнем обмена веществ, значительная часть которого направлена на обеспечение работы натриевого насоса в мембранах и поддержание состояния возбуждения. Химические основыпередачи нервного импульса по аксону уже обсуждались в гл. 5, разд. Б, 3. Последовательное раскрытие сначала натриевых и затем калиевых каналов можно считать твердо установленным. Менее ясным остается вопрос, сопряжено лиизменение ионной проницаемости, необходимое для распространения потенциала действия, с какими-либо особыми ферментативными процессами. Нахманзон указывает, что ацетилхолинэстераза присутствует в высокой концентрации на всем протяжении мембраны нейрона, а не только в синапсах [38, 39]. Он предполагает, что увеличение проницаемости к ионам натрияобусловлено кооперативным связыванием нескольких молекул ацетилхолина смембранными рецепторами, которые либо сами составляют натриевые каналы, либо регулируют степень их открытия. При этом ацетилхолин высвобождается из участков накопления, расположенных на мембране, в результате деполяризации. Собственно, последовательность событий должна быть такова, что изменение электрического поля в мембране индуцирует изменение конформации белков, а это уже приводит к высвобождению ацетилхолина. Под действием аце-тилхолинэстеразы последний быстро распадается, и проницаемость мембраныдля ионов натрия возвращается к исходному уровню. В целом приведенное описание отличается от описанной ранее схемы синаптической передачи только в одном отношении в нейронах ацетилхолин накапливается в связанной с белками форме, тогда как в синапсах — в специальных пузырьках. Существует мнение, что работа калиевых каналов регулируется ионами кальция. Чувствительный к изменению электрического поля Са-связывающий белок высвобождает Са +, который в свою очередь активирует каналы для К" , последнее происходит с некоторым запозданием относительно времени открытия натриевых каналов, что обусловлено различием вконстантах скоростей этих двух процессов [123]. Закрытие калиевых каналов обеспечивается энергией гидролиза АТР. Имеются и другие предположения омеханизмах нервной проводимости [124]. Некоторые из них исходят из того, что нервная проводимость целиком обеспечивается работой натриевого насоса. [c.349] Расстояние между пресинаптической и посГсинаптической мембранами—синаптическая щель — может достигать 15—20 нм. В мионевральном соединении разрыв еще больше— до 50—100 нм. В то же время существуют синапсы с сильно сближенными и даже сливающимися пресинаптической и постсинаптической мембранами. Соответственно реализуются два типа передачи. При больших щелях передача является химической, при тесном контакте возможно прямое электрическое взаимодействие. Здесь мы рассмотрим химическую передачу. [c.382] Выяснив электрические свойства клетки в состоянии покоя, рассмотрим процессы, связанные с возбуждением мембраны. Состояние возбужденияможно определить как временное отклонение мембранного потенциала от потенциала покоя, вызванное внешним стимулом. Этот электрический или химический стимул возбуждает мембрану, изменяя ее ионную проводимость, т. е. сопротивление в контуре снижается (рис. 5.4). Возбуждение распространяется от стимулированного участка к близлежащим областям мембраны, в которыхнаблюдается изменение проводимости, а следовательно, и потенциала. Такое распространение (генерация) возбуждения называется импульсом. Различаются два типа импульсов потенциал действия, когда сигнал распространяется неизмененным от участка возбуждения к нервному окончанию, и локальный потенциал,. быстро уменьшающийся по мере удаления от участка возбуждения. Локальные потенциалы обнаружены в синапсах воз-буждающие постсинаптические потенциалы (е. р. з. р.) и ингибиторные постсинаптические потенциалы ( . р.з.р.)) и в сенсорных нервных окончаниях рецепторные или генераторные потенциалы). Локальные потенциалы могут суммироваться, т. е. они могут увеличиваться при последующих возбуждениях, тогда как потенциалы действия не обладают такой способностью-и возникают по принципу все или ничего . [c.115] Рис. 6. . а — схема нервного волокна с синапсом. Показаны системытранспорта (АТРаза) и три различные системы пассивного транспорта. Справа — хемовозбудимая транспортная система, регулируемая молекулой непроме-диатора, например канал в постсинаптической мембране мышечной концевой пластинки, пропускающий ионы калия и натрия слева — отдельно К а+- и К+-каналы в мембране аксона, управляемые электрическим полем и открываемые при деполяризации бив — проводимость натрия gNг (б) и калня ё к, (в), а также входящий натриевый /ка и выходящий калиевый /к токи после деполяризации (60 мВ). Четко дифференцированная кинетика двух процессов N3 и к подразумевает существование индивидуальных молекулярных структур для пассивного натриевого и калиевого транспорта. [c.131] КИ открытие электрического синапса Фершпаном и Поттером произошло в 1959 г., когда нейронная теория окончательно вытеснила ретикулярную. [c.189] Электрические синапсы сравнительно редки, и их роль в центральной нервной системе высших организмов пока неясна. Фершпан и Поттер открыли их в брюшном нерве краба, а позднее их обнаружили в многочисленных организмах моллюсках,, членистоногих и млекопитающих. В противоположность химическому синапсу, гдепрохождение импульса несколько задерживается из-за высвобождения и диффузии медиатора, сигнал через электрический синапс передается быстро. Физиологическая важность таких синапсов может, следовательно, быть связана с необходимостью быстрого сопряжения специфических клеток. [c.189] Заслуживает внимания еще особенно полезная клеточная-линия — линия клеток РС 12, клонированная из феохромоцитомы — опухоли хромаффинной ткани надпочечника. Клетки РС 12 аналогичны хромаффинным клеткам по их способности синтезировать, запасать и высвобождать катехоламины. Подобно ненейрональным клеткам, они размножаются, но под действием N0 они перестают делиться, участвуют в нейритных процессах и становятся очень похожими насимпатические нейроны. Они приобретают электрическую возбудимость, отвечают на ацетилхолин и даже образуют функциональные холинэргические синапсы. Клетки РС 12 используются в качестве модельных систем для изучениядифференциации нейронов, действия гормональных и трофических факторов, функции и метаболизма гормонального рецептора (см. с. 325). [c.369] Основу каждой НС составляют относительно простые, в большинстве случаев — однотипные элементы (ячейки). Далее под нейроном будет подразумеватьсяискусственный нейрон, то есть ячейка НС (рис. 19.1). Каждый нейрон характеризуется своим текущим состоянием по аналогии с нервными клеткамиголовного мозга, которые могут быть возбуждены или заторможены. Он обладает группой синапсов — однонаправленных входных связей, соединенных с выходами других нейронов, а также имеет аксон — выходную связь данного нейрона, с которой сигнал (возбуждения или торможения) поступает на синапсы следующих нейронов. Каждый синапс характеризуется величиной синаптической связи или ее весом и ,, который по физическому смыслу эквивалентен электрической проводимости. [c.704] Сигналы, проводимые нейронами, передаются от одной клетки к другой в особыхместах контакта, называемых синапсами (рис. 18-3). Обычно эта передача осуществляется, как это ни странно на первый взгляд, непрямым путем. Клетки электрически изолированы друг от друга пресииаптическая клетка отделена от постсинаптической промежутком-синаптической щелью. Изменение электрического потенциала в пресинаптической клетке приводит квысвобождению вещества, называемого ненромедиатором (или нейротрансмиттером), которое диффундирует через синаптическую щель ивызывает изменение электрофизиологического состояния постсинаптической клетки. Та- [c.73]
|
Рис. 18-3. Схема типичного синапса. Элеггричесжий сигнал, приходящий в окончание аксона клетки А, приводит к высвобождению в синаптическую щельхимическое го посредника (иейромеднатораХ который вызывает электрическое изменение в мембране деидрита клетки В. Широкая стрелка указывает направление передачи сигнала, Аксон одного нейрона, такого как изображенный на рис. 18-2, образует иногда тысячи выходных синаптических соединений с другими клетками. И наоборот, нейрон может принимать сигналы через тысячи входных синаптических соединений, находящихся на его дендритах и теле. |
|
 <=""
img="" style="border: none; display: block;
margin-left: auto; margin-right: auto;">
<=""
img="" style="border: none; display: block;
margin-left: auto; margin-right: auto;">Самый простой способ передачи сигнала от нейрона к нейрону-это прямое электрическое взаимодействие через щелевые контакты. Такие электрпескк сшишсы между нейронами встречаются в некоторых участках нервной системы у многих животных, в том числе и у позвоночных. Главное преимущество электрических синапсов состоит в том, что сигнал передается без задержки. С другой стороны, эти синапсы не приспособлены к выполнению некоторых функций и не могут так тонко регулироваться, как химические синапсы, через которые осуществляется большинство связей между нейронами. Электрическая связьчерез щелевые контакты была рассмотрена в главе [c.93] Скелетные мышечные волокна позвоночных, подобно нервным клеткам, способны возбуждаться под действием электрического тока, и нервно-мышечноесоедшенае (рис. 18-24) может служить хорошей моделью химического синапсавообще. На рис. 18-25 сравнивается тонкая структура этого синапса с типичным синапсом между двумя нейронами головного мозга. Двигательный нерв и иннервируемую им мышцу можно отделить от окрузкаюшей ткани и поддерживать вфункционирующем состоянии в среде определенного состава. Возбуждая нерв через наружные электроды, можно с помошью внутриклеточного микроэлектрода регистрировать ответ одиночной мышечной клетки (рис. 18-26). Микроэлектрод сравнительно легко ввести в волокно скелетной мышцы, так как это очень крупная клетка (порядка 100 мкм в диаметре). [c.96] Два простых наблюдения показывают, что для синаптической передачинеобходим приток нонов Са в окончание аксона. Во-первых, если во внеклеточной среде Са отсутствует, медиатор не высвобождается и передачи сигнала не происходит. Во-вторлх, если искусственно ввести Са в цитоплазму нервного окончания при помощи микропипетки, выход нейромедиатора происходит даже без электрической стимуляции аксона, рто трудно осуществить на нервно-мышечном соединении из-за малых размеров окончани аксона поэтому такой эксперимент был проведен на синапсе между гигантскими нейронами кальмара.) Эти наблюдения позволили воссоздать последо вательность событий, происходящих вокончании аксона, которая описана ниже. [c.96]
По́стсинапти́ческий потенциа́л (ПСП) — это вре́менное изменение потенциала постсинаптической мембраны в ответ на сигнал, поступивший с пресинаптического нейрона. Различают:
возбуждающий постсинаптический потенциал (ВПСП), обеспечивающий деполяризацию постсинаптической мембраны, и
тормозный постсинаптический потенциал (ТПСП), обеспечивающий гиперполяризацию постсинаптической мембраны.
ВПСП приближает потенциал клетки к пороговому значению и облегчает возникновение потенциала действия, тогда как ТПСП, напротив, затрудняет возникновение потенциала действия. Условно вероятность запуска потенциала действия можно описать как потенциал покоя + сумма всех возбуждающих постсинаптических потенциалов - сумма всех тормозных постсинаптических потенциалов > порог запуска потенциала действия[1].
Отдельные ПСП обычно невелики по амплитуде и не вызывают потенциалов действия в постсинаптической клетке, однако в отличие от потенциалов действия они градуальны и могут суммироваться. Выделяют два варианта суммации[1]:
временная — объединение пришедших по одному каналу сигналов (при поступлении нового импульса до затухания предшествующего)
пространственная — наложение ВПСП соседних синапсов