

128. Регуляция синтезами секреции гормонов по принципу обратной связи.
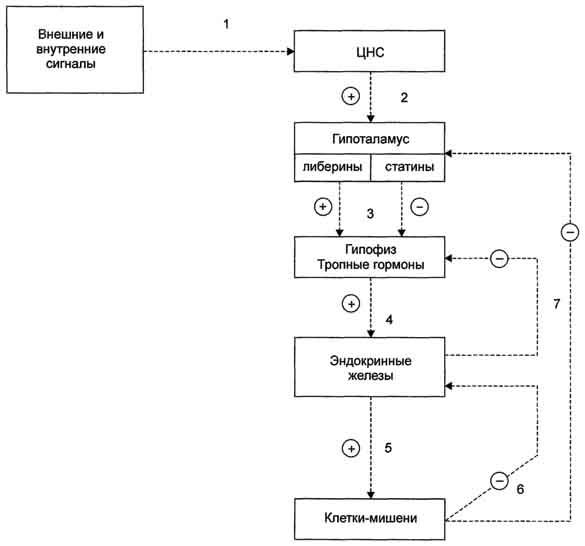
Поддержание уровня гормонов в организме обеспечивает механизм отрицательной обратной связи. Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов, действуя либо на эндокринные железы, либо на гипоталамус. Синтез и секреция тропных гормонов подавляется гормонами эндокринных периферических желёз. Такие петли обратной связи действуют в системах регуляции гормонов надпочечников, щитовидной железы, половых желёз.
Схема взаимосвязи регуляторных систем организма. 1 - синтез и секреция гормонов стимулируется внешними и внутренними сигналами; 2 - сигналы по нейронам поступают в гипоталамус, где стимулируют синтез и секрецию рилизинг-гормо-нов; 3 - рилизинг-гормоны стимулируют (либерины) или ингибируют (статины) синтез и секрецию тройных гормонов.гипофиза; 4 - тройные гормоны стимулируют синтез и секрецию гормонов периферических эндокринных желез; 5 - гормоны эндокринных желез поступают в кровоток и взаимодействуют с клетками-мишенями; 6 - изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов эндокринных желез и гипоталамуса; 7 - синтез и секреция тройных гормонов подавляется гормонами эндокринных желез; ⊕ - стимуляция синтеза и секреции гормонов; ⊝ - подавление синтеза и секреции гормонов (отрицательная обратная связь).
129. Половые гормоны: строение, влияние на обмен веществ и функции половых желез, матки и молочных желез.
Половые гормоны — гормоны стероидной природы, определяющие у человека и животных половую дифференцировку в эмбриональном периоде, характер вторичных половых признаков, функциональную активность репродуктивной системы и формирование специфических поведенческих реакций. Они влияют на многие процессы промежуточного обмена,водно-солевой обмен, а также на состояние адаптационных систем организма. К половым гормонам относятся андрогены, эстрогены и прогестины.
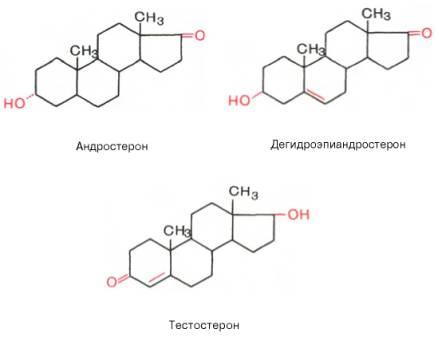
Андрогены — мужские половые гормоны, производные андростана, синтезирующиеся в основном в яичках; некоторое количество андрогенов образуется в коре надпочечников и яичниках. Наиболее активный андроген тестостерон по своей химической структуре является стероидом. Биосинтез андрогенов представляет собой ряд последовательных ферментативных превращенийхолестерина. Основным физиологическим регулятором секреции андрогенов служит лютеинизирующий гормон, взаимодействующий со специфическими циторецепторами. Андрогены, имеющие кетогруппу (СО-группу) при С17, объединяют в группу 17-кетостероидов. В печени андрогены конъюгируются с серной или глюкуроновой кислотами, образуя конъюгаты (парные соединения), которые выводятся с мочой. В крови они содержатся в виде комплексов с липопротеинами, частично в виде свободных глюкуронидов или сульфатов. Тестостерон образуется в яичках, яичниках и надпочечниках. В яичках он продуцируется главным образом клетками Лейдига, в яичниках — тека-клетками овариальных фолликулов, а также в межуточной ткани коркового вещества. В организме взрослого мужчины образуется 4—7 мгтестостерона в сутки, причем около 0,5 мг — в надпочечниках. Яичники и надпочечники взрослой женщины продуцируют примерно 0,5 мгтестостерона в сутки. Основная масса тестостерона, циркулирующего в крови, находится в виде комплекса со специфическим транспортным белком — тестостеронэстрадиолсвязывающим глобулином (ТЭСГ). Связанный с ТЭСГ тестостерон не подвержен метаболическим превращениям. Связывание тестостерона с ТЭСГ служит одним из факторов, определяющих скорость его метаболического клиренса. Метаболические превращения тестостерона осуществляются в печени, почках, кишечнике, легких, коже и других органах. Особое место в его метаболизме принадлежит превращениям в тканях-мишенях. Для метаболического превращения тестостерона в тканях-мишенях характерна 5a-редуктазная реакция, в результате которой образуется 5a-дигидротестостерон. Этот процесс является необходимым этапом биологического действия тестостерона, т.к. именно 5a-дигидроформа связывается с рецепторами тканей-мишеней, 5a-Дигидротестостерон обладает большей андрогенной активностью, чем тестостерон, в связи с чем некоторые исследователи рассматривают тестостерон как прогормон. Биологическое действие тестостерона наиболее специфично в тканях-мишенях, где происходит его избирательное накопление. Рецепторы к тестостерону обнаружены в клетках семенных канальцев, в придатке яичка, предстательной железе, семенных пузырьках, гипоталамусе, матке, фолликулах яичников на определенных стадиях их развития. Андрогенная активность тестостерона проявляется во внутриутробном периоде, когда он, декретируемый яичками плода, обеспечивает половую дифференцировкугипоталамуса, а также формирование внутренних и наружных половых органов по мужскому типу. В период полового созревания под влиянием тестостерона происходит формирование половых органов и развитие вторичных половых признаков. В репродуктивном периоде тестостерон стимулирует определенные этапы сперматогенеза, а также поддерживает половую активность. В женском организме тестостерон оказывает специфическое действие на процессы биосинтеза в клетках матки, а также влияет на развитие фолликулов яичников. Тестостерон обладает выраженным анаболическим действием, связанным со стимуляцией синтеза белка, которое проявляется при формировании фенотипа. Снижение его секреции в мужском организме при гипогонадизме оказывает влияние на формирование наружных половых органов, развитие вторичных половых признаков и сперматогенез. Клиническая симптоматикагипогонадизмаво многом определяется степенью недостаточности выработки тестостерона и тем этапом онтогенеза, на котором это нарушение возникло. У женщин повышенная секреция тестостерона надпочечниками (адреногенитальный синдром, вирилизирующие опухоли надпочечников) или яичниками (вирилизирующие опухоли яичников, склерокистозные яичники) приводит к нарушению генеративной функции яичников, а также к вирилизации.
Эстрогеныявляются производными эстрана, С18-стероидами с ароматическим циклом, фенольной гидроксильной группой при С3 и кетогруппой или гидроксилом при С17. Биосинтез эстрогенов как биохимический процесс представляет собой ароматизацию С19-стероидов, катализируемую комплексом ферментов, локализованных в микросомах. У женщин детородного возраста основная масса эстрогенов синтезируется в яичнике, содержащем зреющий фолликул или желтое тело. Синтез эстрогенов в фолликуле определяется взаимодействием двух стероидпродуцирующих структур зернистого слоя и текаклеток. При этом в последних под регулирующим влиянием лютеинизирующего гормона осуществляется синтез С19-стероидов — андрогенов, которые перемещаются в клетки зернистого слоя, где происходит процесс их ферментативной ароматизации и превращения в эстрогены под контролем фолликулостимулирующего гормона. Синтез эстрогенов в зреющем фолликуле является одним из основных факторов, определяющих функцию гипофизарно-овариальной системы, т.к. повышение концентрации эстрогенов в крови в фазе роста фолликула вызывает преовуляторный выброс лютеинизирующего и фолликулостимулирующего гормонов, которые необходимы для завершения процесса созревания вторичного фолликула и овуляции. Биосинтез эстрогенов путем ароматизации С19-стероидов происходит не только в стероидпродуцирующих железах внутренней секреции, но и во многих тканях организма (жировой ткани, мышцах, печени, почках и др.). В крови эстрогены находятся в основном в виде комплексов с транспортными белками. Образование таких комплексов служит одним из факторов регуляции биологической активности и интенсивности обмена эстрогенов. Основным направлением метаболизма эстрогенов является гидроксилирование стероидного ядра их молекулы в различных положениях. На направленность метаболизма эстрогенов влияет ряд факторов. Так, интенсивность С16-гидроксилирования возрастает при увеличении массы тела, дисфункции печени, снижении концентрациитиреоидных гормоновв крови. Метаболизм эстрогенов происходит в органах-мишенях, почках, коже, эритроцитах и др., однако центральная роль в этом процессе принадлежит печени. Эстрогены, циркулируя в печени, метаболизируются в ней и с желчью попадают в желудочно-кишечный тракт. При этом часть эстрогенов всасывается обратно в кровь, подвергаясь реактивации. В печени образуются водорастворимые конъюгаты эстрогенов и их метаболиты с глюкуроновой и серной кислотами. Кишечно-печеночный цикл и процессы активации — инактивации эстрогенов являются механизмами, регулирующими их обмен и выведение из организма. Нарушение этих механизмов объясняет появление гиперэстрогении у мужчин, больных циррозом печени. Эстрогены и их метаболиты экскретируются с мочой и калом. Физиологический эффект эстрогенов определяется их взаимодействием с рецепторами клеток-мишеней. Рецепторы эстрогенкомпетентных клеток обладают неодинаковым сродством к различным природным и синтетическим эстрогенам. Так, связывание эстрадиола выше, чем синэстрола (гексэстрадиола), эстрона, эстриола (в порядке убывания), что соответствует биологической активности перечисленных эстрогенов в отношении клеток-мишеней. Основное биологическое действие эстрогенов заключается в их влиянии на формирование и функционирование женских половых органов. Эстрогены вызывают увеличение матки за счет роста стромы миометрия и эндометрия, под влиянием эстрогенов осуществляется васкуляризация эндометрия и рост его желез. На протяжении менструального цикла под влиянием меняющегося уровня секреции эстрогенов происходят морфологические изменения слизистой оболочки матки и эпителия влагалища. Под контролем эстрогенов находятся некоторые ключевые этапы яичникового цикла: они дифференцированно влияют на чувствительность клеток зернистого слоя и текаклеток к лютеинизирующему и фолликулостимулирующему гормонам. Эстрогены участвуют в формировании вторичных половых признаков, оказывают модулирующее влияние на различные структуры гипоталамуса, что, в частности, находит выражение в формировании специфического полового поведения. Важную роль играют эстрогены в регуляции функции молочных желез также на жировой обмен, обмен веществ в костной ткани и коже, систему мононуклеарных фагоцитов.

Прогестины.Желтым телом яичника, корой надпочечников, яичками и плацентой синтезируется стероидный гормон прогестерон, который относится к С21 стероидам. Образование прогестерона в яичниках вне беременности регулируется лютеинизирующим гормоном, а при беременности — хорионическим гонадотропином. Механизм действия прогестерона такой же, как у всех стероидных гормонов. Этот гормон взаимодействует со специфическими цитоплазматическими белковыми рецепторами, образуя комплекс, который переносится в ядро клетки и активирует определенные структуры хроматина. В результате стимулируется синтез специфических белков и изменяется функциональное состояние органов-мишеней. Прогестерон участвует в регуляции циклической трансформации эндометрия. Под его влиянием происходят секреторное преобразование эндометрия в лютеиновой фазе менструального цикла, а также функциональные изменения в маточных трубах, влагалище и эпителии молочных желез. Одной из основных физиологических функций прогестерона является торможение сократительной функции миометрия, особенно во время беременности. Снижение секреции прогестерона желтым телом (вне беременности) приводит к недостаточности лютеиновой фазы менструального цикла и к неполноценной секреторной трансформации эндометрия.