

86. Метилирование днк. Представление о метилировании чужеродных и лекарственных соединений.
Метилирование ДНК— это модификация молекулыДНКбез изменения самой нуклеотидной последовательности ДНК, что можно рассматривать как частьэпигенетическойсоставляющейгенома. МетилированиеДНК заключается в присоединенииметильной группыкцитозинув позиции С5 цитозинового кольца.
У человека за процесс метилирования ДНК отвечают три фермента, называемые ДНК-метилтрансферазами1, 3a и 3b (DNMT1, DNMT3a, DNMT3b), соответственно. Предполагается, что DNMT3a и DNMT3b — это de novo метилтрансферазы, которые осуществляют формирование паттерна метилирования ДНК на ранних стадиях развития, а также его изменения в процессе дифференцировки клеток. Существует гипотеза о том, что метилирование ДНК de novo вызывается, в частности,интерферирующими РНКпри помощи РНК-зависимого метилирования ДНК — процесса, возникшего в ходе эволюции с целью репрессии мобильных элементов генома. DNMT1 является ДНК-метилтрансферазой, которая поддерживает метилированное состояние ДНК, присоединяя метильные группы к одной из цепей ДНК в точках, где другая, комплементарная ей цепь, метилирована. Белок DNMT3L гомологичен другим DNMT-белкам, но не имеет каталитической активности. Вместо этого, DNMT3L поддерживает de novo метилтрансферазы, способствуя связыванию этих ферментов с ДНК и стимулируя их активность.
Метилирование. Ворганизмеметилированиюмогут подвергатьсяамины,фенолыитиолы. В результатеметилированияобразуются соответствующие N-, О- и S-метильные конъюгаты. Приметилированиичужеродных соединений и некоторыхметаболитовпереносчиком метильных групп являетсякоферментS-аденозилметионин. С участием метильных групп этогокоферментапроисходитметилированиеперечисленных выше соединений.Реакцииметилированияпроисходят под влиянием ферментных систем (метилтрансфераз).
Η-метилирование. При N-метилировании метильная группа S-аденозилметионина под влиянием N-метилтрансферазы присоединяется катомамазотаметаболитовили чужеродных соединений.
О-Метилирование. Этому типу конъюгации подвергаются соединения, содержащие фенольные группы. Под влияниемферментов(О-метилтрансфераз) метильная группакоферментаS-аденозилметионина присоединяется катомамкислородафенольныхгидроксилов. Дляреакцииметилированияфеноловкромекоферментатребуется присутствиеионовмагнияилиионовдругих двухвалентныхметаллов. Соединения, содержащие одну фенольную группу, при наличии указанныхферментовне метилируются.
S- метилирование. Некоторые чужеродные соединения, содержащие тиоловые группы (-SH), ворганизмеподвергаютсяметилированию. При этом метильная группакоферментаS-аденозилметионина в присутствииферментов(метилтрансфераз) переносится катомамсерыметаболитовили чужеродных соединений с образованием соответствующих S-метилпроизводных этих соединений.
87. Источники и образование одноуглеродных групп. Тетрагидрофолиевая кислота и цианкобаламин и их роль в процессах трансметилирования.
Образование и использование одноуглеродных фрагментов.Особое значение реакций катаболизма серина и глицина заключается в том, что они сопровождаются образованием одноуглеродного метиленового фрагмента (-СН2-). Метиленовая группа в молекуле метилен- Н4-фолата может превращаться в другие одноуглеродные группы (фрагменты): метенильную (-СН=), формильную (-НС=О), метильную (-СН3) и формиминогруппу (-CH=NH). Ещё один источник формального и форми-мино-фрагментов - гистидин. Катаболизм гистидина происходит только в печени (очень небольшой процент в коже) в результате следующих реакций

Конечными продуктами катаболизма гистидина являются глутамат, NH3 и одноуглеродные фрагменты - формимино-Н4-фолат и формил-Н4-фолат.
Все образующиеся производные Н4-фолата играют роль промежуточных переносчиков и служат донорами одноуглеродных фрагментов при синтезе некоторых соединений: пуриновых оснований и тимидиловой кислоты (необходимых для синтеза ДНК и РНК), регенерации метионина, синтезе различных формиминопроизводных (формиминоглицина и т.д.)
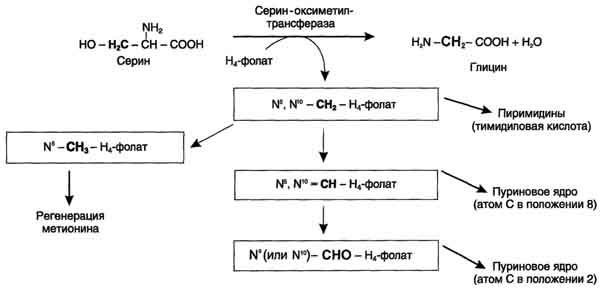
Перенос одноуглеродных фрагментов к акцептору необходим не только для синтеза ряда соединений, но и для регенерации свободного Н4-фолата в печени.
Доказано, что коферментные функции ТГФК непосредственно связаны с переносом одноуглеродных групп, первичными источниками которых в организмеявляются β-углеродныйатомсерина, α-углеродныйатомглицина,углеродметальных группметионина,холина, 2-й углеродныйатоминдольного кольцатриптофана, 2-й углеродныйатомимидазольного кольцагистидина, а такжеформальдегид,муравьиная кислотаиметанол. К настоящему времени открыто шесть одноуглеродных групп, включающихся в разнообразные биохимические превращения в составе ТГФК: формильная (—СНО), метильная (—СН3), метиленовая (—СН2—), метенильная (—СН=), оксиметильная (—СН2ОН) и формими-новая (—CH=NH). Выяснено, что присоединение этих фрагментов к ТГФК является ферментативнойреакциейковалентного связывания их с 5-м или 10-матомомазота(или с обоимиатомамивместе). Имеются данные, что производные ТГФК участвуют в переносе одно-углеродных фрагментов прибиосинтеземетионинаитимина(перенос метильной группы),серина(перенос оксиметильной группы), образовании пуриновыхнуклеотидов(перенос формильной группы) и т.д. Недостаточность фолиевой кислоты у человека возникает редко. Гиповитаминоз фолиевой кислоты приводит к нарушению обмена одноуглеродных фрагментов. Такое же нарушение наблюдается и при недостаточности витамина В12, использование которого связано с обменом фолиевой кислоты. Первое проявление дефицита фолиевой кислоты - мегалобластная (макроцитарная) анемия. Она характеризуется уменьшением количества эритроцитов, снижением содержания в них гемоглобина, что вызывает увеличение размера эритроцитов. Причина этих симптомов - нарушение синтеза ДНК и РНК из-за недостатка их предшественников - тимидиловой кислоты и пуриновых нуклеотидов вследствие дефицита производных Н4-фолата. Клетки кроветворной ткани быстро делятся, поэтому они в первую очередь реагируют на нарушение синтеза нуклеиновых кислот снижением скорости эритропоэза. Мегалобластная анемия возникает чаще всего в результате недостаточности фолиевой кислоты и/или витамина В12.