

27.Изменение активности ферментов при болезнях. Наследственные энзимопатии. Происхождение ферментов крови и значение их определения при болезнях.
В основе многих заболеваний лежат нарушения функционирования ферментов в клетке - энзимопатии. Различают первичные (наследственные) и вторичные (приобретённые) энзимопатии. Приобретённые энзимопатии, как и вообще протеинопатии, по-видимому, наблюдают при всех болезнях.
При первичных энзимопатияхдефектные ферменты наследуются, в основном, по аутосомнорецессивному типу. Гетерозиготы, чаще всего, не имеют фенотипических отклонений. Первичные энзимопатии обычно относят к метаболическим болезням, так как происходит нарушение определённых метаболических путей. При этом развитие заболевания может протекать
по одному из ниже перечисленных "сценариев". Рассмотрим условную схему метаболического пути:
![]()
Вещество А в результате последовательных ферментативных реакций превращается в продукт Р. При наследственной недостаточности какого-либо фермента, например фермента Е3, возможны разные нарушения метаболических путей:
Нарушение образования конечных продуктов. Недостаток конечного продукта этого метаболического пути (Р) (при отсутствии альтернативных путей синтеза) может приводить к развитию клинических симптомов, характерных для данного заболевания:
![]()
Клинические проявления. В качестве примера можно рассмотреть альбинизм. При альбинизме нарушен синтез в меланоцитах пигментов - меланинов. Меланин находится в коже, волосах, радужке, пигментном эпителии сетчатки глаза и влияет на их окраску. При альбинизме наблюдают слабую пигментацию кожи, светлые волосы, красноватый цвет радужки глаза из-за просвечивающих капилляров. Проявление альбинизма связано с недостаточностью фермента тирозингидроксилазы (тирозиназы) - одного из ферментов, катализирующего метаболический путь образования меланинов
Накопление субстратов-предшественников. При недостаточности фермента Е3 будут накапливаться вещество С, а также во многих случаях и предшествующие соединения. Увеличение субстратов-предшественников дефектного фермента - ведущее звено развития многих заболеваний:
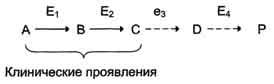
Клинические проявления. Известно заболевание алкапгонурия, при котором нарушено окисление гомогентизиновой кислоты в тканях (гомогентизиновая кислота - промежуточный метаболит катаболизма тирозина). У таких больных наблюдают недостаточность фермента окисления гомогентизиновой кислоты - диоксигеназы гомогентизиновой кислоты, приводящей к развитию заболевания. В результате увеличиваются концентрация гомогентизиновой кислоты и выведение её с мочой. В присутствии кислорода гомогентизиновая кислота превращается в соединение чёрного цвета - алкаптон. Поэтому моча таких больных на воздухе окрашивается в чёрный цвет. Алкаптон также образуется и в биологических жидкостях, оседая в тканях, коже, сухожилиях, суставах. При значительных отложениях алкаптона в суставах нарушается их подвижность.
Нарушение образования конечных продуктов и накопление субстратов предшественников. Отмечают заболевания, когда одновременно недостаток продукта и накопление исходного субстрата вызывают клинические проявления.
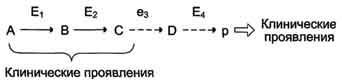
Клинические проявления. Например, у людей с болезнью Гирке (гликогеноз I типа) наблюдают снижение концентрации глюкозы в крови (гипогликемия) в перерывах между приёмами пищи. Это связано с нарушением распада гликогена в печени и выходом из неё глюкозы вследствие дефекта фермента глюкозо-6-фосфатфосфатазы. Одновременно у таких людей увеличиваются размеры печени (гепатомегалия) вследствие накопления в ней не используемого гликогена.
Особый интерес для клиники представляет исследование активностииндикаторныхферментоввсыворотке крови, так как по появлению вплазмеилисыворотке кровиряда тканевыхферментовв повышенных количествах можно судить о функциональном состоянии и поражении различных органов (например,печени, сердечной и скелетной мускулатуры). При остром инфаркте миокарда особенно важно исследоватьактивностькреатинкиназы, АсАТ, ЛДГ и оксибутиратдегидрогеназы. При заболеванияхпечени, в частности при вирусном гепатите (болезнь Боткина), всыворотке кровизначительно увеличиваетсяактивностьАлАТ и АсАТ, сорбитолдегидрогеназы,глутаматдегидрогеназыи некоторых другихферментов. озрастаниеактивности ферментовсыворотки кровипри многих патологических процессах объясняется прежде всего двумя причинами: 1) выходом в кровяное руслоферментовиз поврежденных участков органов илитканейна фоне продолжающегося ихбиосинтезав поврежденныхтканях; 2) одновременным повышением каталитическойактивностинекоторыхферментов, переходящих вкровь. Возможно, что повышениеактивности ферментовпри «поломке» механизмов внутриклеточной регуляцииобмена веществсвязано с прекращением действия соответствующих регуляторов иингибиторов ферментов, изменением под влиянием различных факторов строения и структурымакромолекулферментов.
28.Применение ферментов для лечения болезней. Применение ферментов как аналитических реагентов при лабораторной диагностике (определении глюкозы, этанола, мочевой кислоты и т.д.). Иммобилизованные ферменты
Второе направление медицинской энзимологии – энзимодиагностика– развивается по двум путям. Один путь – использованиеферментовв качестве избирательныхреагентовдля открытия и количественного определения нормальных или аномальных химическихвеществвсыворотке крови,моче, желудочном соке и др. (например, выявление при помощиферментовглюкозы,белкаили другихвеществвмоче, в норме не обнаруживаемых). Другой путь – открытие и количественное определение самихферментовв биологическихжидкостяхпри патологии. Оказалось, что рядферментовпоявляется всыворотке кровипри распадеклеток(отсюда их название «некротическиеферменты»). Для диагностики органических и функциональных поражений органов итканейшироко применяются отдельные ферментныетесты, выгодно отличающиеся от других химических диагностическихтестов, используемых в клинике, высокой чувствительностью испецифичностью. Известно около 20тестов, основанных на количественномопределении активности ферментов(иизоферментов), главным образом вкрови(реже вмоче), а также в биоптатах (кусочкитканей, полученные прибиопсии). Следует отметить, что из огромного числаферментов(более 3500), открытых в природе (частично и ворганизмечеловека), в диагностической энзимологии используется лишь ограниченный наборферментови для весьма небольшого числа болезней (гепатиты, инфаркт миокарда, органические поражения почек, поджелудочной железы,печении др.). Так, уровеньлипазы,амилазы,трипсинаихимотрипсинавкровирезко увеличен присахарном диабете, злокачественных поражениях поджелудочной железы, болезняхпечении др. Резко повышается всыворотке кровиуровень двух аминотрансфераз, креатинкиназы (и ее изоформ) илактатдегидрогеназы(и ее изоформ) при инфаркте миокарда; умеренно повышено их содержание при пораженияхтканеймозга ипечени. Определяют, кроме того,активностькислой фосфатазы(уровень повышен прикарциномепредстательной железы), щелочнойфосфатазы, холинэсте-разы и некоторых других органоспецифическихферментов(например, гистидазы, уроканиназы, глицинамидинотрансферазы) всыворотке кровипри патологиикостной ткани,печени, метастатическихкарциномахи т. д. Доказано, что органы итканичеловека характеризуются специфическим ферментным и изоферментным спектром, подверженным не только индивидуальным, но и суточным колебаниям. Существует большой градиентконцентрацииферментовмежду внутриклеточными и внеклеточными частями тела. Поэтому любые, даже незначительные, поврежденияклеток(иногда функциональные расстройства) приводят к выделениюферментовво внеклеточное пространство, откуда они поступают вкровь. Механизм гиперферментации (повышенное содержаниеферментоввкрови) до конца не расшифрован. Повышение уровня внутриклеточныхферментоввплазме кровипрямо зависит от природы повреждающего воздействия, времени действия и степени повреждения биомембранклетоки субклеточных структур органов. В оценке ферментныхтестовдля диагностических целей особое значение имеет знание периода полужизни (полураспада) вплазме кровикаждого из диагностическихферментов, что делает важным выбор точного времени для ферментного анализакрови. Весьма существенным является также знание особенностей распределения (топографии)ферментовв индивидуальных органах итканях, а также их внутриклеточной локализации.
В последнее время стали применять ферментырестрикции– специфическиеэндонуклеазыкатализирующие разрывы межнуклео-тидных связейДНК, для диагностикифенилкетонурии, α- и β-талассемии и другихнаследственныхболезней человека. Метод основан наполиморфизмерестрикционных фрагментовДНК.
Из представленных данных следует, что диагностическая энзимология может служить основой не только для постановки правильного и своевременного диагноза болезни, но и для проверки эффективности применяемого метода лечения.
Дальнейшее развитие диагностической энзимологии преимущественно идет по двум перспективным направлениям медицинской энзимологии: по пути упрощения и рациональной модификации уже испытанных методов и по пути поиска новых органоспецифических (тканеспецифических) ферментовиизоферментов.
Третье направление медицинской энзимологии – энзимотерапия, т.е. использованиеферментови модуляторов (активаторовиингибиторов) действияферментовв качествелекарственных средств, имеет пока небольшую историю. Досихпор работы в этом направлении почти не выходят за рамки эксперимента. Исключение составляют некоторые про-теиназы:пепсин,трипсин,химотрипсини их смеси (абомин,химопсин), которые применяют для лечения ряда болезней пищеварительного тракта. Помимо протеиназ, ряд другихферментов, в частностиРНКаза, ДНКаза, гиалуронидаза,коллагеназы, эластазы, отдельно или в смеси с протеина-зами используются при ожогах, для обработки ран, воспалительных очагов, устранения отеков, гематом, келоидных рубцов, кавернозных процессов при туберкулезе легких и др.Ферментыприменяются также для лечения сердечно-сосудистых заболеваний,растворениясгустковкрови. В нашей стране разработан первый в мире препарат иммобилизованной стрептокина-зы, рекомендованный для лечения инфаркта миокарда. Калликреины –ферментыкининовой системы используются для снижения кровяногодавления.
Важной и многообещающей областью энзимотерапии является применение ингибиторов ферментов. Так, естественныеингибиторыпротеиназ (α1-трипсин, α1-химотрипсин, α-макроглобулин) нашли применение в терапии острых панкреатитов, артритов, аллергических заболеваний, при которых отмечается активацияпротеолизаифибринолиза, сопровождающаяся образованием вазоактивныхкининов.
В последнее время получило признание применение в онкологической клинике ферментовбактериальной природы в качествелекарственных средств. Широко используется L-аспарагиназа (выпускается в промышленных количествах и L-глутамин(аспарагин)азадля лечения острых и хронических формлейкозовилимфогранулематозов. Более десятка описанных в литературе бактериальныхферментовиспытаны в основном на животных с перевивнымиопухолямиили на раковыхклеткахопухолейчеловека и животных, выращенных вкультуре ткани. Основными постулатамиприменения ферментовв онкологии являются различия вметаболизмеклетокопухолейпо сравнению с обменом в нормальной, здоровой,клетке. В частности, современные стратегия и тактика энзимотерапии опухолевых поражений учитывают разную чувствительность нормальных и опухолевыхклетокк недостатку (дефициту) незаменимых (так называемых эссенциальных) факторов роста. К таким ростстимулирующим факторам относятся не только пищевые факторы (витамины, незаменимыеаминокислоты, макро- имикроэлементы), но и ряд так называемых заменимыхвеществ, включая заменимыеаминокислоты, к недостатку которых опухолеваяклеткаоказывается в силу особенностей ее обмена более чувствительной, чем нормальная. Лечебный эффект, например, L-аспарагиназы и L-глутамин (аспа-рагин)азыприлейкозах, вероятнее всего, объясняется необратимым распадом какглутамина, так иаспарагина. Оказалось, что опухолевыеклеткидля своего роста и размножения нуждаются ваминокислотахизорганизма, поскольку сами лишены способности синтезировать амидыаминокислот, в то время как нормальныеклеткинаделены этой способностью. Был сделан вывод о том, что амидныйазотглутаминаиаспарагинавыполняет вклеткахряд уникальных функций, которые лучше выяснены для глута-мина. В частности, амидныйазотглутаминаоказался абсолютно необходимым и не заменимым другимиаминокислотамиисточникоматомаазотаминимум в 10реакцияхсинтеза, например, пуриновых и пиримидиновыхнуклеотидов, соответственноДНКиРНК,АТФ, ге-ксозаминов,гистидинаи др. Таким образом, не лишенаоснованиягипотеза, что любойферментили агент, катализирующий необратимое расщепление незаменимого для опухолевойклеткипищевого фактора (включаяаминокислоты), может в принципе быть применен в энзимотерапииопухолей, если будут устранены ограничения, связанные с белковойприродой фермента. В оценке эффективностиферментовв экспериментальной и клинической онкологии имеется немало противоречий и очень много пробелов. Положительные результаты, отмеченные в ряде случаев, вселяют надежду, что приготовление стандартных ферментных препаратов (включая создание иммобилизованных форм) в промышленных масштабах и их разумное применение в клинике, организованное на строгой научной основе, несомненно дадут в руки врачей еще одно ценное оружие в борьбе с опухолевыми заболеваниями человека.
Идея применения ферментовв качествелекарственных средств(фармакологииферментов) всегда казалась заманчивой. Однако их нестабильность, короткий период полураспада, нежелательные антигенные свойства, связанные с белковойприродой ферментови опасностью развития аллергическихреакций, трудности доставки к пораженным органам итканям(мишеням) существенно ограничивали возможности использования ферментных препаратов. В разработке методовиммобилизацииферментовнаметились конкретные пути преодоления указанных трудностей: применение водорастворимых, биосовместимыхносителей, например полимолочнойкислоты(легко разлагается ворганизме), использование методов химической модификации имикрокапсулирования, приготовление моно- и поликлональныхантители ферментсодержащих липосом и т.д.
В последнее время интенсивно разрабатываются методы направленного транспорта ферментов, заключенных в своеобразные микроконтейнеры (тениэритроцитов,липосомыи др.), к внешней поверхности которых могут быть прикреплены адресные (векторные) белковыемолекулы(например,иммуноглобулины–антителапротив специфических компонентов органа или ткани-мишени, в частностиопухоли).Иммобилизованные ферментыв качествелекарственных средствначали применять в специальных колонках для экстракорпоральной перфузиикрови(типа искусственной почки). Такое лечение полностью исключает нежелательные воздействия наорганизмчужеродногобелкаи может проводиться длительное время.